Страница
3
Подобная техника была использована в экспериментах Бензера, Реди, Рубина, Толимсона, Цирупски и их коллег, которые исследовали дифференцировку нейронов и сопутствующих (опорных) клеток глаза. Глаз дрозофилы состоит из кристаллоподобного набора повторяющихся единиц, называемых омматидиями (рис. 4A), каждый из которых содержит 8 фоторецепторов (R1-R8). Первая клетка, которая определяет начало дифференцировки в каждом омматидии, является одним из фоторецепторов, R8. Клетки R8 появляются хаотично в области нейроэпителия. Как только началась дифференцировка клетки R8, это приводит к ингибированию дифференцировки ее соседей в клетки R8. Затем дифферениировке подвергаются клетки R2 и R5, после чего R4, R1 и R6 и, наконец, R7 (рис. 4В). Были обнаружены две мутантные линии, в которых глаза развиваются нормально, за исключением того, что не образуется R7 (рис. 4В). Такие линии были названы sevenless (sev--) и brideof-sevenless (boss-- ) для обозначение отсутствия образования R7. При детальном исследовании этих мутантных линий впервые были изучены молекулярные механизмы того, как индукционные взаимодействия между клетками могут влиять на дальнейшее развитие клеток. Ген sevenless кодирует рецептор (называемый Sevenless, или Sev), для которого продукт экспрессии bride-of-sevenless гена, названный Boss, является лигандом. Во время развития R8 индуцирует образование R7. Это происходит, когда Boss, экспрессируемый на поверхности клетки R8, связывается с Sev, который расположен на клетке-предшественнице R7 (рис. 4С). Взаимодействие между Boss и Sev активирует внутриклеточный домен Sev рецептора, тирозинкиназу, которая инициирует сигнальный каскад в клетке R7, приводящий к ее дифференцировке. Сигнальный каскад является сложным путем, который включает в себя последовательную активацию серии протеинкиназ (ферментов, которые фосфорилируют белки), что приводит к ингибированию негативного влияния, а также увеличению позитивного влияния на экспрессию генов R7. Большое количество сигналов, которые приводят к изменениям в эспрессии генов, осуществляют это через тирозинкиназы, которые управляют данными клеточными каскадами.
Происхождение клеток в ЦНС млекопитающих
Изучать последовательность развития клеток в ЦНС млекопитающих технически более сложно, потому что отдельные клетки труднее идентифицировать и заполнить красителем. Удачными методиками исследования здесь показали себя картирование путей развития генетически помеченных клеток у химерных животных — эмбрионов и взрослых, а также инфицирование клеток ЦНС развивающихся животных специально созданными вирусами (рис. 5). Подобные вирусы сконструированы таким образом, что они перманентно включаются в состав хромосом клетки-хозяина, реплицируются во время деления клетки, и, следовательно, передаются потомкам этой клетки. Таким образом, информация, находящаяся в вирусе, не уменьшается при делении клеток. Присутствие вируса может быть затем обнаружено на любой стадии развития клеток, используя белок, который он кодирует. При условии, что изначально количество инфицированных клеток мало, в дальнейшем можно будет сделать вывод, что кластер клеток, содержащих данный белок, является клоном, потомством той самой клетки, которая была ранее инфицирована.
Например, когда такой вирус был введен в глаз новорожденного крысенка, а сетчатка была исследована во взрослом возрасте, белок содержали как глиальные клетки, так и несколько типов нейронов. Таким образом, общая клетка-предшественница в сетчатке при делении образует как нейрональные, так и глиальные клетки.
В отличии от пиявки или дрозофилы, в сетчатке грызунов нет специфической последовательности развития клеток, когда одна клетка образует несколько различных типов нейронов. В данном случае, вероятно, клетка--предшественник имеет некоторую внутреннюю информацию (intrinsic competence) о том, как отвечать на внешние сигналы и сигналы, которые находятся в окружающей ее среде. Эти сигналы меняются со временем, что приводит к последовательному формированию клеток разных типов. С другой стороны, когда подобные эксперименты проводились в коре больших полушарий, клоны, которые содержали бы и глиальные клетки, и нейроны, были довольно редки. Это дает основания полагать, что в момент инфицирования отдельные популяции клетокпредшественииков глиальных и нервных клеток уже сформировались в вентрикулярной области коры. Более того, клоны большей частью содержали исключительно пирамидные или непирамидные клетки, показывая, что разделение между этими двумя линиями происходит на раннем этапе нейрогенеза.
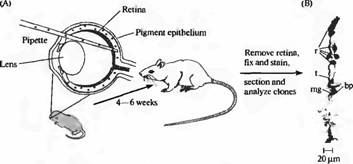
Рис. 5. Клетки одного клона помечены при помощи введения ретровирусных маркеров в сетчатку крысы. (А) Ретровирус, кодирующий бета-галактозидазу был введен в глаз между сетчаткой и пигментным эпителием в раннем периоде развития, что привело к заражению нескольких предшественников клеток сетчатки. (В) Окрашивание сетчатки взрослого при помощи гистохимической реакции на бета галактозидазу выявляет кластеры, содержащие метку и являющиеся потомками одной клетки-предшественницы. Изображение клона включает в себя 5 палочек (г), одну биполярную клетку (bp) находящуюся вблизи к терминали палочки (t)
Резюмируя вышесказанное, в нервной системе простых организмов возможный спектр дальнейшего развития клетки (lineage history) ограничивает потенциал ее развития. В ЦНС более сложных животных существуют индукционные взаимодействия между клетками, которые играют исключительную роль в направлении развития клетки. В хорошо известной и искрометной аналогии Сидней Бреннер охарактеризовал два пути развития клетки как «план по-американски» и «план по-европейски»: «по-европейски» то, кто ты есть (какой нейрон), определяется твоими предками; «по-американски» это определяется твоими соседями.