Страница
2
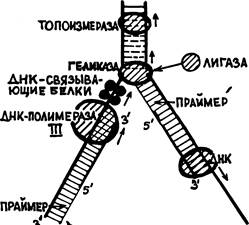
Между геликазой и ДНК-полимеразой III образуется некоторый участок обнаженной 1-й нити. 2-я нить тоже еще ничем не прикрыта. Эти две нити после ухода геликазы могут вновь сомкнуться. Чтобы этого не происходило, вплотную за геликазой на 1-ю нить садятся четыре, так называемых «ДНК-связывающих белка». Им не приписывают иных функций, кроме защиты от восстановления двойной спирали ДНК близ вершины вилки .
Пройдя до вершины вилки разошедшихся нитей материнской ДНК тандем геликаза-ДНК-связывающие белки — ДНК-полимераза III останавливается (см. рис. 28). Топоизомераза уходит дальше по двухнитевой материнской ДНК, а геликаза разрывает сахаро-фосфатную связь на 2-й нити. Уплотненные на участке, прилегающем к вилке, витки двойной спирали расправляются, 1-я нить ДНК вместе с сидящими на ней белками вращается вокруг своей оси, а вокруг этой нити поворачивается и отрезанный кусок 2-й нити, временно связанный с геликазой. Этот кусок называют «фрагментом Оказаки» — по имени ученого, обнаружившего появление таких фрагментов при редупликации. После снятия напряжения нити двойной спирали материнской ДНК снова могут начать расходиться. Но до этого с отрезанного конца фрагмента Оказаки другая праймаза начинает на нем построение нового рибонуклеотидного праймера. Затем геликаза освобождает фрагмент и уходит вперед, а специальный фермент «лигаза» пришивает начало фрагмента Оказаки на прежнее место — ко 2-й нити материнской ДНК. Заметим, что лигаза (М=96 тыс.) в клетке E.coli представлена наиболее многочисленной популяцией — около 200 молекул. Из чего следует, что она выполняет не случайные «ремонтные» работы, а является полноправным членом совокупности ферментов, обеспечивающих редупликацию ДНК (подобно значению ниток для хирурга).
Когда праймер готов, впереди него, по направлению к 5'-концу 2-й материнской нити ДНК садится ДНК-полимераза I. Начинается строительство нити, комплементарной к этому фрагменту 2-й нити, опять в направлении 3'— 5', считая по этой нити. ДНК-полимераза I доходит до конца фрагмента Оказаки и снимается. Этим заканчивается 1-й этап редупликации (рис. 28).
Между тем праймер, оставшийся в начале 1-й нити разрушается некой «рибонуклеазой Н», — ферментом, рвущим нить РНК, находящуюся в комплексе с нитью ДНК. На его место ДНК-полимераза II ставит «правильные» дезоксирибонуклеотиды. В то же время топоизомераза, геликаза, а за ними и ДНК-полимераза III продвигаются вперед.
Начинается 2-й этап редупликации. Вилка репликации тоже продвигается, прилегающий к ней участок материнской двунитевой ДНК уплотняется и весь синтезирующий тандем останавливается. Геликаза опять разрезает 2-ю нить, образуя второй фрагмент Оказаки. Так же, как раньше, на обрезанном (временно) конце фрагмента создается праймер, к нему «подсаживается» ДНК-полимераза I и начинает копировать второй фрагмент Оказаки, т.е. 2-ю нить материнской ДНК. Отличие второго этапа будет только в том, что на пути этой полимеразы встретится праймер, оставшийся от копирования 1-го фрагмента Оказаки. Но ДНК-полимераза I, в отличие от всех прочих ДНК-полимераз, обладает еще и 5'—3' экзонуклеазной активностью, т. е. в направлении своего движения. Она разрушает праймер и доходит до того места, с которого начинала копирование 1-го фрагмента Оказаки ее предшественница. Остается только связать фосфодиэфирной связью эти два куска новосинтезированной комплементарной нити. Естественно, что это делает вездесущая ДНК-лигаза.
![]()
![]()
![]()
![]() рибонуклеаза Н
рибонуклеаза Н
ДНК-полимеразаII
Рис. 28
Тем временем в районе образования уже третьей вершины вилки редупликации происходят точно такие же события, как на 2-м этапе редупликации. Скорость этого процесса оценивается как, примерно, 1000 нуклеотидов в секунду у бактерий 100 — у животных и 20 — у растений.
Весьма вероятно, что в то же самое время аналогичные процессы расплетания двойной спирали с образованием фрагментов Оказаки и комплементарного построения новых нитей ДНК идут и с противоположного конца материнской ДНК. Разумеется, там ДНК-полимераза III непрерывно двигается вдоль той нити ДНК, которую мы назвали 2-й, а на фрагменты Оказаки разрезается 1-я нить. Когда два движения встречаются, две дочерние копии исходной ДНК оказываются готовы. (Их «сошьет» все та же лига-за.) Кстати оказалось, что длина фрагментов Оказаки у E.coli (1—2 тысячи нуклеотидов) значительно больше, чем у эукариотов (меньше 200). Не лишено интереса совпадение этой последней цифры с длиной ДНК в нуклеосоме (см. ниже).
Более сложная модель движения репликативной вилки предполагает формирование реплисомы – мультиферментного комплекса более высокого уровня организации. Этот комплекс состоит из функционального праймосомо-праймазного комплекса, геликазы, полимеразы III, и, возможно, гиразы. Такой комплекс может обеспечивать удлинение лидирующей цепи и одновременно инициацию праймерной РНК, а также достраивание ДНК при синтезе отстающей цепи. Две реплисомы, работающие согласованно в двух вилках репликации, которые движутся в противоположных направлениях вдоль кольцевой хромосомы, сделали бы эту модель еще более изящной.
Репликация кольцевых дуплексов. Репликация инициируется также в точке начала репликации (ori).
Растущие цепи образуют репликативные вилки, пересещающиеся либо в двух (вверху), либо в одном (внизу) направлении в зависимости от природы точки начала репликации.
В некоторых кольцевых геномах в каждой цепи имеется своя точка начала репликации (например, в митохондриальной ДНК животных ). Синтез одной цепи начинается в точке oriR. Когда новая цепь доходит до точки oriD, начинается синтез другой цепи. Синтез инициируется путем образования праймерной РНК.
Некоторые двуцепочечные кольцевые хромосомы реплицируются альтернативным способом, называемым репликацией по типу катящегося кольца. В этом случае двуцепочечная кольцевая ДНК надрезается специфическим ферментом в уникальном сайте одной цепи (точке начала катящегося кольца), и к образовавшемуся в результате надреза 3΄ - гидроксильному кольцу с помощью полимеразы III присоединяются нуклеотиды; при этом матрицей служит интактная замкнутая цепь. Таким образом в вилке синтезируется только лидирующая нить. По мере синтезалидирующей цепи происходит вытеснение 5΄-конца надрезанного кольца как одиночной цепи. В результате длина лидирующей цепи может превышать длину матрицы в 2-5 раз. Такой способ репликации используют фаги М13 или фX174 (их зрелые геномы одноцепочечные кольцевые ДНК)на поздних стадиях инфекционного процесса, после того как инфицирующая ДНК превращается в двуцепочечную кольцевую форму. Постоянно отделяющиеся одиночные цепи ДНК, образуемые при репликации по типу катящегося кольца, надрезаются в каждой точке начала репликации и замыкаются с образованием зрелых форм, упаковываемых в вирусные частицы. Фаг λ использует такой способ репликации при образовании двуцепочечной линейной вирусной ДНК. Субстратной матрицей в этом случае является двуцепочечная кольцевая ДНК, которая была реплицирована после превращения на ранних этапах инфекции линейной вирусной ДНК в кольцевую репликативную форму.