Страница
4
Таким образом, для биосинтеза одной молекулы сахарозы необходимы 3 макроэргические фосфатные связи, эта реакция необратима.
В нефотосинтезирующих тканях некоторых растений, например, в корнеплодах сахарной свеклы, клубнях картофеля и других сахароза может образоваться из свободной фруктозы:
УДФ-глюкоза + фруктоза ↔ сахароза + УДФ.
Реакция катализируется ферментом сахарозо-УДФ-глюкозилтрансферазой и в зависимости от условий может быть направлена как в сторону синтеза, так и в сторону расщепления сахарозы.
Синтез крахмала. Крахмал состоит из амилозы, представляющей собой неразветвленную цепь остатков ос-глюкозы, соединенных α(1-4)-связями, и амилопектина — разветвленной молекулы, в которой наряду с α(1-4)- имеются и α(1-6)-связи. В строении этих двух сахаров, входящих в крахмал, имеются существенные различия, поэтому механизмы их биосинтеза различны.
Донором глюкозных остатков при биосинтезе амилозы может служить уридиндифосфатглюкоза-(УДФК). Для ее образования в реакционной среде необходимо наличие затравки, в качестве которой могут служить полисахариды, построенные всего лишь из 3—4 остатков глюкозы, соединенных α(1-4)-связями.
Остатки глюкозы переносятся на акцептор (затравку), где и происходит удлинение цепи. Реакция идет по схеме:
УДФГ + акцептор (Г)к —— УДФ + акцептор (Г)к + 1,
где Г — остатки глюкозы.
Фермент, катализирующий эту реакцию, называется УДФГ-крахмалглюкозилтрансферазой.
У большинства растений активным донором глюкозы является не УДФ-глюкоза, а аденозиндифосфатглкжозα(АДФГ). Реакция присоединения глюкозных остатков от АДФГ к низкомолекулярному акцептору идет аналогичным путем и катализируется ферментом АДФГ-крахмал-глюкозилтрансферазой.
Синтез разветвленной молекулы амилопектина, имеющей α (1-6)-связи, происходит при помощи фермента α-глюкантрансферазы (Q-фермент).
В синтезе крахмала участвуют и Д-фермент или глюкозилтрансфераза, образующий α(1-4)-связи и участвующий в образовании затравки.
Распад крахмала происходит при участии двух процессов - гидролиза и фосфоролиза.
Гидролитический распад крахмала осуществляется под действием четырех ферментов класса гидролиз α-амилаза, катализирует расщепление α(1-4)-связи, причем связи разрываются беспорядочно. Конечный продукт такого распада — мальтоза, глюкоза, декстрины. Под действием β-амилазы происходит расщепление α (1-4)-связей с образованием остатков мальтозы. Фермент глюкоамилазы катализирует последовательное отщепление остатков глюкозы от молекулы крахмала. Амилопектин-1,6-глюкозидаза или R-фермент катализирует расщепление α(1-6)-связей в молекуле амилопектина, т. е. действует на точки ветвления.
Фосфоролиз — это присоединение фосфорной кислоты по месту разрыва глюкозидной связи между остатками моносахаридов в цепи полисахарида, при этом происходит образование глюкозо-1-фосфата. Эта реакция катализируется ферментомаглюконфосфорилазой, относящимся к классу трансфераз. Крахмал в растении может подвергаться очень быстрому распаду, так как ферменты распада находятся во всех органах растения.
Синтез целлюлозы. Целлюлоза построена из остатков β-глюкозы. В биосинтезе целлюлозы принимает участие не свободная глюкоза, а ее ГДФ-производное — гуанозиндифосфатглюкоза при участии фермента целлюлозосинтетазы по схеме:
ГДФ — глюкоза + (глюкоза) к→ ГДФ + (глюкоза)к + 1
Распад целлюлозы идет преимущественно гидролитическим путем под действием фермента целлюлазы до дисахарида целлобиозы.
Транспорт углеводов осуществляется в виде сахарозы. В процессе фотосинтеза образуется много углеводов, и в этой связи большое значение имеет отток ассимилятов в другие части клетки из хлоропластов. Проникновение через мембрану хлоропластов фосфорилированных гексоз и сахарозы затруднено, наиболее легко через мембраны хлоропластов проникают триозофосфаты (ФГА и ФДА). Предполагается, что образующиеся сложные углеводы распадаются на триозофосфаты и в таком виде передвигаются в цитоплазму, где могут служить материалом для ресинтеза гексоз, сахарозы, крахмала.
Межклеточный паренхимный транспорт осуществляется двумя путями — по плазмодесмам (симпласту) или по свободному пространству (аппопласту). Сахароза, образовавшаяся в клетках мезофилла листа, десорбируется в аппопласт. Выходя из паренхимных клеток в аппопласт, сахароза расщепляется инвертазой на гексозы. Гексозы передвигаются по аппопласту к передаточным клеткам проводящих пучков по градиенту концентраций. При соприкосновении с передаточными клетками флоэмы они снова превращаются в сахарозу. Далее происходит загрузка ситовидных трубок, сахароза поступает против градиента концентраций, и требуется расход энергии (АТФ).
Предполагается, что сахароза преодолевает мембрану с помощью переносчика в комплексе с протоном. При этом благодаря работе Н+-АТФ-азы ионы Н+ выкачиваются из клеток флоэмы, а затем поступают обратно по градиенту рН, увлекая за собой сахарозу против градиента ее концентрации. Основной транспортной формой углеводов по флоэме служит сахароза (С12Н22О11). У некоторых видов наряду с сахарозой транспортной формой углеводов служат олигосахара (рафиноза, стахиоза), а также некоторые спирты.
85. Дыхание как совокупность последовательных окислительно-восстановительных процессов
В процессе дыхания участвует сложная цепь окислительно-восстановительных превращений углеводов и жиров. Под окислением какого-либо соединения понимают процесс потери им электрона (протона), под восстановлением — их присоединения, протекающие по одному из следующих путей.
Непосредственное присоединение к веществу кислорода, например, в процессе образования воды:
2Н2 + O2 → 2Н2О.
Нередко процесс отдачи электрона может происходить без участия кислорода:
![]()
Одновалентная медь служит донором электрона и окисляется до Сu2+. В обратной реакции Сu2+ — акцептор электронов восстанавливается до Сu .
Перенос электронов во многих окислительно-восстановительных реакциях сопровождается одновременным переносом протонов, как при окислении янтарной кислоты в фумаровую:

Для непрерывного протекания этой реакции необходимо наличие в системе акцептора водорода.
Способность вещества к окислению сильно возрастает, если оно предварительно гидратируется:
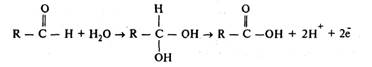
В живых системах окисление одного соединения всегда сопряжено с восстановлением другого. Любая окислительно-восстановительная система может функционировать как окислитель или как восстановитель в зависимости от соотношения величины ее собственного потенциала и потенциала системы, с которой она взаимодействует. Максимальную величину потенциала в клетке имеет кислород, благодаря чему он служит универсальным окислителем.