Страница
3
У большинства видов папоротников листья совмещают 2 функции - фотосинтеза и спороношения (рис. 2), однако у многих видов наблюдается диморфизм листьев - одни выполняют функцию фотосинтеза, а другие - только спороношения, например у страусника, трихоманеса. У некоторых видов папоротников (у ужовниковых, у осмунды) имеется диморфизм частей листа, при котором одна часть выполняет функцию фотосинтеза, а другая часть - спороношения. Палеонтологический материал показывает, что все 3 типа листьев существовали уже в раннем палеозое и формировались независимо друг от друга.
Ход развития корней папоротников до сего времени мало изучен. Как уже отмечалось, при формировании зародыша зародышевый корень закладывается одновременно со стеблем и листом, при этом смещается гаусторией вбок. В дальнейшем у одних видов он развивается одновременно с листом, а у других видов несколько задерживается в росте и появляется позднее, поэтому производит впечатление придаточного корня.
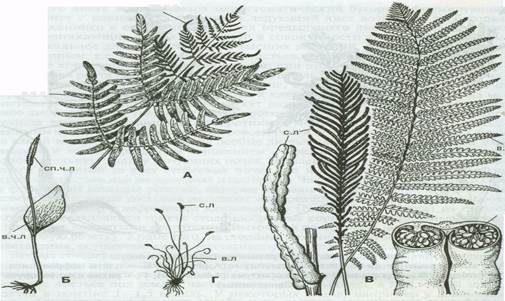
Рис. 2 - Спороносные листья папоротников: А - осмунды (Оsmunda); Б - ужовника(Оphioglossum); В - страусника (Маtteuccia); Г - трихоманеса (Тrichomanes); Д - схизеи (Schizaeae); в. ч. л - вегетативная часть листа; сп. ч. л - спороносная часть листа; в. л - вегетативный лист; с.л - спороносный лист
У одних видов корни могут появиться на поверхности побега в год его заложения, а у других видов - лишь через 1-2 года. От настоящих придаточных корней семенных растений корни папоротников (так же, как у плаунов и хвощей) отличаются тем, что не могут закладываться на уже сформированных частях побегов. Корни папоротников живут 3-4 года. У некоторых видов, например у нефролеписа, корни, загибаясь вверх, способны превращаться в олиственные побеги.
Анализ вегетативных органов папоротников выявляет способность превращения одного органа в другой, т.е. указывает на то, что дифференциация на органы у них не всегда жестко закреплена генетически. Это находится в прямой связи с древностью папоротников. Сходство анатомического строения стеблей, черешков листьев и корней также указывает на единство происхождения всех вегетативных органов.
У большинства современных видов папоротников стебли имеют диктиостелу, однако ход морфогенеза у разных видов различен. У многих изученных папоротников стебель молодого растения построен по типу протостелы, затем формируется эктофлоическая или амфифлоическая сифоностела, а с началом образования листовых лакун происходит формирование диктиостелы. Она представляет собой цилиндр, в центре которого расположена паренхима сердцевины, затем следуют внутренняя флоэма, ксилема и внешняя флоэма, пронизанные паренхимой листовых лакун. На поперечном срезе листовые лакуны представлены сердцевинными лучами, а между ними по кольцу располагаются концентрические проводящие пучки с ксилемой в центре и флоэмой по периферии. При мацерации все живые ткани сердцевины, сердцевинных лучей и флоэмы разрушаются и остается лишь ксилема в виде сетчатого цилиндра, отсюда и название диктиостела - сетчатая стела. Флоэма одночленная, состоит только из ситовидных клеток. Ксилема сложена в основном лестничными, частично кольчатыми трахеидами. Наряду с диктиостелой у некоторых древних видов на протяжении всей жизни сохраняется протостела (у лигодиума, глейхении), эктофлоическая сифоностела (у циатеи) или амфифлоическая сифоностела (у видов глейхении, у диптериса). Все это позволяет сделать вывод о направленности эволюции стелы от протостелы через сифоностелу к диктиостеле, что подтверждается палеоботаническим материалом. У пермских папоротников описана протостелическая структура, у триасовых - в основном сифоностелическая, у большинства современных - диктиостела, при которой осуществляется наибольший контакт проводящих тканей с живой паренхимой, а следовательно, улучшается водоснабжение. Размножение папоротников осуществляется в основном за счет спор. Большинство видов папоротников - равноспоровые растения; количество разноспоровых видов невелико. Для всех папоротников характерно отсутствие специализированных спороносных побегов - стробилов. У большинства папоротников спорангии сгруппированы в сорусы; у мараттиевых, срастаясь между собой, они образуют синангии (рис. 3). У наиболее примитивных видов одиночные спорангии располагаются по краям листьев либо на вершинах их лопастей, при этом каждый спорангий снабжается самостоятельной жилкой. Это весьма напоминает верхушечное расположение спорангиев на концах васкуляризованных теломов у риниофитов.

Рис. 3 - Типы размещения спорангиев и сорусов: А - расположение одиночных спорангиев: 1 - верхушечное по отношению к долям листа у даваллиевых (Davalliacеае); 2 - краевое у трихоманеса (Тrichomanes); Б - расположение сорусов: 1 - краевое у гименофилла (Нуmophillum); 2 - на нижней стороне листа у кочедыжника ломкого (Суstopteris fragilis); В - синангии у мараттиевых (Магаttiacеае): 1 - Аngiopteris; 2 - Магattiа; 3 - Сhristensiа; Г -перемещение спорангиев на нижнюю поверхность листа в ходе морфогенеза у схизеи (Schizaea); сп -спорангий
У большинства же папоротников спорангии или сорусы располагаются на нижней поверхности листьев. Чаще всего они приурочены к главной и боковым жилкам, реже (например, у орляка) - близ края листа. Расположение спорангиев на нижней стороне листа оказывается биологически выгодным: во-первых, обеспечивается надежная защита спорангиев во время их созревания и одновременно не снижается интенсивность фотосинтеза. Во-вторых, обеспечивается более равномерное рассеивание спор, в треть их создается возможность большего продуцирования спорангиев на всей поверхности листа, чем только по краю. Ход морфогенеза спороносных листьев у некоторых видов рода схизеи показывает возможный путь перемещения спорагниев на нижнюю поверхность листа в ходе филогенеза. На ранних этапах развития листа схизеи спорангии закладываются по его краям, но в результате более быстрого развития тканей верхней стороны листа происходит загибание его краёв вниз. В результате оказываются на нижней стороне листа. Место прикрепления спорангии к листу называется плацентой. В ходе эволюции происходило разрастание плаценты, и она приобретала продолговатую или шаровидную форму, что увеличивало площадь возможного размещения спорангиев. Кроме того, на плаценте спорангии располагаются в определенной последовательности, a следовательно, и созревание и вскрывание их происходит неодновременно. Растянутость созревания спор во времени гарантирует более надежное их распространение. Увеличение количества спорангиев на плаценте достигается также увеличением длины ножки спорангия. Разрастание плаценты и увеличение длины ножки коррелятивно не связаны, эти два процесса происходили в разных систематических группах независимо друг от друга.
Большое биологическое значение имеет защита сорусов в период их созревания. В простейшем случае сорусы прикрываются загибающимся вниз краем листа. У более специализированных видов формируется особое покрывало индузиум, возникающее в результате локального разрастания плаценты либо поверхностных тканей листа. По характеру заложения и строению стенки спорангия папоротники подразделяются на лепто- и эуспорангиатн ы е. У эуспорангиатных папоротников спорангий возникает из группы клеток и имеет многослойную стенку, у лептоспорангиатных он возникает из одной клетки и имеет однослойную стенку. И тот и другой тип спорангиев обнаружены у древних папоротников. Механизмы вскрывания спорангиев также чрезвычайно многообразны. В простейшем случае на вершине спорангия имеется небольшой участок сильно утолщенных клеток - пора. При подсыхании спорангия происходит его разрыв на контакте тонкостенных и толстостенных клеток. Однако уже у древних папоротников возникло так называемое кольцо - полоска из толстостенных клеток. У представителей одних семейств оно занимает горизонтальное положение, у других - косое, у третьих - вертикальное. Примитивным считается кольцо сплошное, или замкнутое; более совершенным - неполное, при котором часть клеток кольца остается неутолщенным (так называемое устье); по нему происходит разрыв стенки. Примером может служить спорангий щитовника мужского (Driopteris filix-mas), имеющий форму двояковыпуклой линзы и расположенный на длинной ножке. Однослойная стенка спорангия состоит из крупных тонкостенных клеток. По гребню спорангия, начиная от ножки, проходит кольцо, состоящее на 2/3 из толстостенных клеток, а 1/3 приходится на устье. Клетки кольца имеют утолщения на трех стенках - на двух радиальных и на внутренней - тангентальной. При подсыхании клетки кольца теряют воду, что приводит к уменьшению их объема. Большая сила сцепления втягивает тонкую внешнюю тангентальную стенку внутрь клетки, и она прогибается, притягивая при этом радиальные стенки друг к другу. Это приводит к уменьшению длины окружности кольца и создает большое натяжение в области устья. Происходит