Отдел красные водоросли
Среди криптонемиальных, как и среди немалиальных, встречаются формы не только одноосевого, но и многоосевого строения.
Порядок гигартинальные—Gigartinales
Порядок характеризуется тем, что ауксилярные клетки дифференцируются до оплодотворения в непосредственной близости от карпогона, так что прокарпий имеется; в качестве ауксилярной функционирует клетка, участвующая в формировании таллома, а не специальная — добавочная. Размножение можно рассмотреть на примере рода анфельция. Зрелые мужские и женские гаметофиты представлены вертикальными, в различной степени разветвленными многоосевыми талломами, вырастающими от хорошо развитых базальных корочек. На мужских талломах в плодущих участках зрелые антеридии образуют один ряд вытянутых клеток. При помещении живого материала в морскую воду под микроскопом можно наблюдать, как вытянутые спермации с силой отбрасываются от таллома на значительное расстояние. На женских талломах плодущие участки находятся вблизи верхушки. Прокарпий Ahnfeltia concinna состоит из крупной поддерживающей клетки и трехклеточной карпогонной ветви, верхняя клетка которой представляет собой карпогон. Поддерживающая клетка функционирует как ауксилярная. После оплодотворения карпогона и слияния его с поддерживающей клеткой, одновременно являющейся ауксилярной, развиваются гонимобласты — карпоспорофиты, врастающие в ткань сердцевины таллома, где они могут сохраняться дольше года до того, как освободятся карпоспоры.
При прорастании карпоспоры образуется корковидный таллом, морфологически отличный от гаметангиальных талломов-гаметофитов в виде вертикальных разветвленных или неразвегвленных пластинок многоосевого строения, растущих от базальных корочек. Приблизительно через три недели после прорастания карпоспор на корковидном талломе начинается формирование тетраспорангиев, в которых развиваются крестообразно расположенные тетраспоры. Тетраспоры прорастают как карпоспоры, образуя сначала корочки, на которых, однако, возникают многоосевые талломы анфельции, сходные с встречающимися в природе. Корочки, развившиеся из тетраспор, никогда не образуют тетраспорангиев, а корочки из карпоспор никогда не дают вертикальных талломов анфельции. Таким образом, у Ahnfeltia concinna гетероморфная смена генераций. У ряда представителей был продемонстрирован жизненный цикл Polysiphonia-типа.
У ряда видов из класса флоридеофициевых было описано аномальное сочетание репродуктивных ортанов; среди этих сообщений многие касаются представителей порядка гигартинальных. Например, у Cystodiniwn purpureum, в норме характеризующегося изоморфной сменой поколений и морфологически сходными гаметофитами и тетраспорофитами, у некоторых растений на одном и том же талломе присутствовали и тстраспорангии, и цистокарпии.
В ряде случаев у одного и того же вида обнаруживаются разные циклы развития. Например, у Gigartina siellata наблюдается или половой цикл с PetraceUis cruenta в качестве тетраспорофита, или же бесполый цикл, по-видимому, с апогамным развитием цистокарпиев.
Порядок родимениальные — Rhodymeniales
Порядок объединяет формы многоосевого строения. Ауксиляр-ные клетки отчленяются от клетки, получившейся в результате деления несущей клетки до оплодотворения карпогона. Ауксилярные клетки — добавочные, специальные клетки, не принимающие участия в построении вегетативного таллома. Есть прокарпии, в состав которых в одних случаях, кроме карпогона, входит одна ауксилярная клетка, в других — две. У одних форм тетраспорангии с тетраэдрическим расположением тетраспор, у других — с крестообразным. Представителем служит род ломен-гария, имеющий трубчатый, полый с перетяжками или без них таллом с боковым ветвлением.
До сравнительно недавнего времени к этому порядку относили широко распространенную по берегам Европы гюдоросль родимению пальчатую {Rhodymenia palmata). Таллом многоосевого типа имеет вид листа, сидящего на суженном основании. В настоящее время эта водоросль под названием пальмария пальчатая выделена в новый порядок пальмари-альные главным образом на основе строения тетраспо-рангиев, с характерными клетками-ножками. Тетраспорангиальный зачаток формируется из коровой клетки таллома, которая при этом увеличивается, претерпевает митоз и неравновеликий цитокинез с образованием крупного тетраспорангия и более мелкой клетки-ножки. Ядро спорангия делится мейотически, вслед за чем содержимое спорангия распадается на четыре крестообразно расположенные тетраспоры. Тетраспоры содержат в цитоплазме везикулы, возможно с клеющим материалом, способствующим прикреплению освободившихся тетраспор к субстрату. После освобождения тетраспор клетка-ножка увеличивается и делится с образованием новой клетки-ножки и нового тетраспорангия. Долгое время женский гаметофит был неизвестен, наблюдались лишь мужские индивиды, образующие мелкие сорусы антеридиев по поверхности таллома. Однако в 1980 г. в лабораторной культуре P. palmata, полученной из тетраспор этой водоросли, собранных вдоль берегов Канады, был обнаружен женский микроскопический гаметофит. Таким образом, мужские и женские гаметофиты P. palmata морфологически различаются. После оплодотворения тетраспорофит начинает развиваться на женском гаметофите непосредственно из зиготы, из тетраспор образуются гаплоидные гаметофиты — мужские и женские.
Порядок церамиальные—Ceramiales
Этот порядок, наиболее эволюционно продвинутый и богатый видами, характеризуется одноосевым строением таллома, наличием прокарпия и тем, что ауксилярные клетки дифференцируются только после того, как произошло оплодотворение карпогона. Карпогонная ветвь всегда четырехклеточная, неразветвленная, несущая клетка—перицентральная, т. е. всегда отчленяется от клетки центральной оси плодущей ветви. После оплодотворения ауксилярная клетка отделяется от несущей карпогонную ветвь клетки.
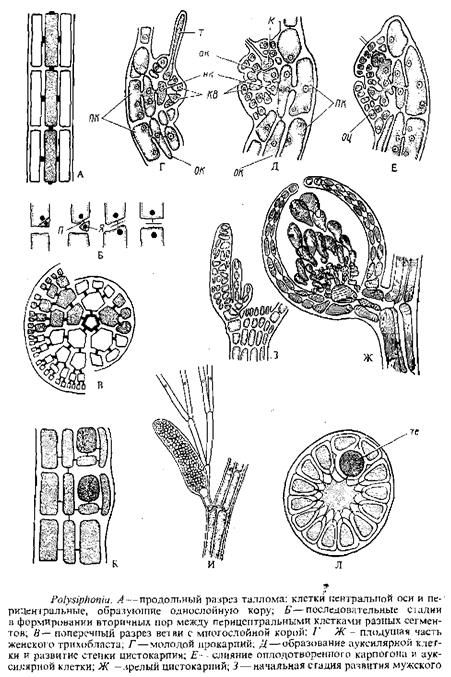
Как было недавно показано для Callithamnion cordatum, необходимая предпосылка образования ауксилярной клетки после оплодотворения— контакт карпогона и клетки, несущей карпогонную ветвь, от которой отчленяется ауксилярная клетка. Без такого контакта ауксилярная клетка не образуется, что позволяет предположить участие в ее формировании одного или нескольких экзогенных гормонов. Образование ауксилярных клеток наблюдалось приблизительно через 40 ч, а диплоидизация их — через 72 ч после соединения в культуре мужского и женского гаметофитов.
Брюшко оплодотворенного карпогона или непосредственно сливается с рядом расположенной ауксилярной клеткой, или слияние происходит через посредство коротких выростов.
В качестве примера лучше всего рассмотреть широко распространенный в южных и северных морях род полисифонии, который еще в начале XX в. был подробно изучен цитологически и с точки зрения чередования морфологических фаз. Как указывалось, жизненный цикл этой водоросли получил название Polysiphonia-тип и позднее был обнаружен у многих представителей вышестоящих порядков флоридеофици-евых. Талломы полисифонии представлены темно-малиновыми разветвленными кустиками, ветви которых увенчаны верхушечными клетками. Верхушечная клетка посредством нескольких косых стенок отчленяет клетки-сегменты. Эти клетки на стороне, где их высота больше, отделяют обычно инициальную клетку боковой ветви еще до деления сегмента на центральную и перицентральную клетки. Инициальная клетка боковой ветви развивается или в моносифоновые, состоящие из одного ряда клеток ветвящиеся трихо-дласты, или же дает начало боковым ветвям, повторяющим строение главной оси. После этого клетка-сегмент претерпевает последовательные деления продольными перегородками, которые отчленяют периферические клетки от центральной. Перицентральные клетки связаны с центральной посредством первичных пор, а между перицентральными клетками соседних сегментов образуются вторичные поры. При этом на морфологически нижнем конце перицентральной клетки отшнуровывается маленькая клетка с одним ядром, естественно соединенная порой с клеткой, от которой отделилась. Эта маленькая клеточка сливается с соответствующей перицентральной клеткой ниже расположенного сегмента. В результате две друг над другом расположенные перицентральные клетки оказываются соединенными порой. У некоторых видов перицентральные клетки делятся с образованием многослойной коры.