Отдел красные водоросли
У вышестоящих порядков красных водорослей ауксилярные клетки формируются рядом с карпогоном до оплодотворения карпогона или только после того, как оно произошло. Совокупность ауксилярной клетки с карпогоном называется прокарпием. Здесь нет необходимости в образовании длинных ообластемных нитей, связывающих карпогоп и ауксилярные клетки, ауксилярная клетка просто сливается с брюшком оплодотворенного карпогона, после чего из нее развиваются гонимобласты с карпоспорами. Карпоспорангии часто располагаются тесными группами — цисто-карпиями, которые у многих представителей одеты псевдопаренхиматозной оболочкой, развивающейся из соседних с карпогоном клеток.
Жизненные циклы красных водорослей отличаются большим разнообразием, что в последнее время было продемонстрировано на основе следующих данных, полученных разными методами. Наблюдения в природе дают представление о встречаемости морфологических фаз и репродуктивных органов, на них развивающихся. В условиях лабораторной культуры можно проследить прорастание репродуктивных клеток и развитие из них талломов до состояния, когда они дают репродуктивные органы, т. е. в культуре можно строго установить последовательность морфологических фаз. Особенно плодотворным оказался метод лабораторных культур для установления связей между гаметангиальной и тетраспорангиальной фазами, различающимися морфологически и долгое время рассматриваемыми как самостоятельные таксоны. Цитологический метод позволяет установить число хромосом каждой морфологической фазы, место в жизненном цикле сингамии, мейоза и других цитологических деталей. Ниже приводятся жизненные циклы, обоснованные совокупностью вышеперечисленных данных,
У красных водорослей наиболее распространен жизненный цикл, состоящий в последовательности гаметангиальной, карпоспорангиальной и тетраспорангиальной фаз, причем первая и последняя морфологически сходны, а карпогониальная фаза развивается на гаметангиальной. Такая изоморфная смена гаметофитов и тетраспорофитов впервые была описана для Polysiphonia violaceae. Поэтому такой жизненный цикл, присущий большинству красных водорослей, обычно называют Polysiphonia-тип. Его можно встретить помимо порядка церамиальных, куда относится род полисифония, и у представителей порядков криптонемиальные, гигартинальные, родимениальные и гелидиальные.
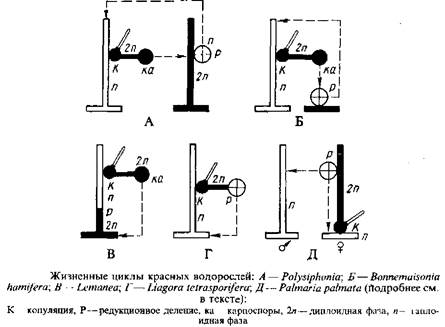
Предположение о жизненном цикле Polysiphonia-типа у представителей вышестоящих порядков класса флоридеофициевых в последние годы подтверждено строгими культуральными экспериментами.
Цикл развития Bonnemaisonia hamifera-тина, который можно назвать гетероморфной сменой генераций, характеризуется последовательностью гаметанги-альной, карпоспорапгиальной и тетраспорангиальной фаз, причем все три фазы морфологически неодинаковы. Карпогониальная фаза развивается на гаметангиальной. В последние годы такой жизненный цикл был описан не только для многих представителей порядка немалиальных, но и для криптонемиальных и гигартинальньгх.
У Liagora tctrasporifera в жизненном цикле чередуются гаметангиальная и тетраспорангиальная фазы, морфологически различающиеся, причем тетраспорангиальная фаза развивается на гаметангиальной; мейоз осуществляется в тетраспорангиях, карпоспорофит отсутствует.
У Palmaria paimata также отсутствует карпоспорофит и наблюдается чередование гаметангиальной и тетраспорангиальной фаз, причем вторая развивается на первой. Однако женский гаметофит здесь крохотный, а мужской — крупный, морфологически неотличимый от тетраспорофита.
Приведенные жизненные циклы характеризуются спорической редукцией, так как мейоз происходит при образовании тетраспор.
Жизненный цикл, описанный пока только для двух родов — леманеа и батрахоспермум, отличается соматической редукцией. В апикальной клетке предростка, развивающегося из диплоидных карпоспор, происходит мейоз и из нее вырастает гаплоидная гаметангиальная фаза.
Сказанным не исчерпывается все разнообразие жизненных циклов красных водорослей.
Красные водоросли делятся на два класса: бангиофициевые и флоридеофициевые
КЛАСС БАНГИОФИЦИЕВЫЕ — BANGIOPHYCEAE
Клетки часто со звездчатым хроматофором, заключающим пиреноид. Поры между клетками, как правило, отсутствуют. Кар-погон без трихогины, после оплодотворения содержимого карпогона непосредственно делится с образованием карпоспор. Бесполое размножение обычно моноспорами.
Представителями могут служить роды порфира и бангия. Род порфира характеризуется листоватыми талломами истинно паренхиматозного строения, прикрепленными своими основаниями к субстрату и достигающими 50 см в длину, редко более. Пластинка состоит из одного слоя клеток или двухслойная. Клетки содержат по одной или по две пластиды. Пластинки у разных видов неодинаковой формы с цельными или зазубренными краями, различной окраски и толщины. Проростки сначала представлены однорядными нитями. Последовательные продольные деления клеток этих нитей приводят к образованию листовидных талломов. Порфира распространена как в южных, так и в северных морях в прибрежной литоральной зоне.
Род бангия, включающий как морские, так и пресноводные виды, имеет таллом, представленный неразветвленными нитями, вначале прикрепленными к субстрату базальной клеткой; с возрастом от нижних клеток развиваются песептированные ризоиды. Нити сначала однорядные, но в более старых участках клетки претерпевают продольные радиальные деления, так что на поперечном срезе таллома видны сектора.
Зрелые талломы порфиры и бангии размножаются половым путем. У порфиры при образовании антеридиев клетки талломов делятся во взаимно перпендикулярных направлениях на ряд мелких клеток, каждая из которых образует по одному спермацию. При разбухании интерцеллюлярной слизи спермации выдавливаются и освобождаются. Карпогоны лишь незначительно отличаются от вегетативных клеток. Типичная трихогина отсутствует. После оплодотворения зигота непосредственно делится на 2—32 карпоспоры, которые освобождаются так же. как и спермации,— благодаря набуханию межклеточной слизи. Сходным образом осуществляется половое размножение бангии. Так. Е. Мань утверждает, что талломы бангии гаплоидны и представляют собой гаметофиты. Они двудомны: мужские талломы продуцируют спермации, которые копулируют с клетками женских нитей, функционирующими как каргюгоны. В результате возникает диплоидная зигота, которая делится на карпоспоры.
Цикл развития. Цикл развития порфиры и бангии интенсивно изучался в последние десятилетия как в природе, так и в искусственных культурах. Для многих видов порфиры и бангии в их жизненном цикле были установлены Conchocelis-стадии: карпоспоры, сначала голые, выделяют оболочку и прорастают униполярно в нитчатые растеньица, образующие на поверхности и внутри известковых раковинок моллюсков розовые пятна1. Эта стадия долгое время считалась самостоятельной водорослью Conchoceiis rosea. Нити конхоцелиса сложены из удлиненных клеток с париетальным хроматофором. Ультраструктура клеток типична для красных водорослей из класса флоридеофициевых: слоистая клеточная оболочка; одиночные тилакоиды в хлоропластах, окруженных двухмембранной хлоропластной оболочкой: периферические тилакоиды: пиреноид, в который проникают одиночные тилакоиды; рассеянные между тилакоидами фибриллы ДНК; в цитоплазме — гранулы багрянкового крахмала. Между соседними клетками Conchocelis-нитей имеются закупоренные пробками поры, сходные с таковыми в клетках флоридеофициевых.