Страница
3
Род хромулина, как и охромопас, представлен одиночными голыми клетками, но с одним жгутиком.
Род уроглена (Uroglena). Виды этого рода обычны в пресноводном планктоне озер и прудов, представлены шаровидными или эллипсоидальными слизистыми колониями, в которых протопласты находятся на концах слизистых.
Семейство динобриевые объединяет свободно живущие или прикрепленные, одиночные или колониальные формы, клетки которых заключены в домик – мертвое, выделяемое протопластом образование. Типичный представитель – род динобрион, виды которого широко распространены в пресноводном планктоне и при массовом развитии нередко вызывают цветение воды в озерах и прудах. Голая, одетая цитоплазматичес кой мембраной клетка заключена в бокаловидном домике, образованном целлюлозными микрофибриллами с петлеобразным расположением; базальньгй конус домика состоит из продольно ориентированных микрофибрилл. В состав домика входит и большое количество аминокислот. Протопласт прикреплен к одной стороне основания домика базальной ножкой, способной к удлинению, сокращению или закручиванию, что позволяет клетке двигаться внутри домика: в этой части клетки находятся многочисленные микротрубочки. Клетка овальных очертаний содержит одно ядро, расположенное между хлоропластами. Два хроматофора различаются по длине, меньший располагается сзади. В хлоропласте можно видеть тилакоиды, жировые капли. На переднем конце более крупного хроматофора имеется глазок, состоящий из рядов электронно-плотных гранул несколько различной величины. В цитоплазме находятся эндоплазматический ретикулум, полирибосомы, митохондрии, в передней трети клетки - одна крупная диктиосома из приблизительно 12 диктиосомальных цистерн, от которых по краям отшнуровываются многочисленные пузырьки. В различных местах цитоплазмы содержатся свободные липидные гранулы, в задней части клетки – крупная вакуоль с хризоламинарином. Клетка имеет два жгутика: длинный и более короткий. Проксимальная часть короткого гладкого жгута несет вздутие, расположенное в непосредственной близости от стигмы. От базального тела к ядерной оболочке тянется поперечно исчерченный корень, кроме того, имеются 7 микротрубчатых корней, распространяющихся на периферию клетки. Клеточное тело соприкасается со стенкой домика не только базальной ножкой, но и сравнительно широкой экваториальной областью, в которой в молодых и делящихся клетках наблюдаются обильные пузырьки – дериваты аппарата Гольджи. Это заставляет предположить, что материал домка секретируется именно этими пузырьками. При продольном делении клетки один из дочерних протопластов или оба покидают материнский домик и, оседая у его края, вращаясь вокруг своей оси, формируют новый домик. Таким образом, возникают древовидные колонии различной формы. При образовании статоспоры базальная ножка, по-видимому, отделяется от стенки домика, и клетка мигрирует к его отверстию, где округляется, теряет жгутики и образует цисту или статоспору. По крайней мере часть статоспор Dinobryon cylindricum – результат гологамного полового процесса.
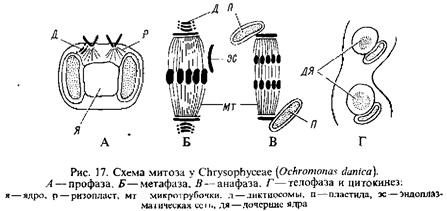
Семейство Синуровые – Synuraceae включает одноклеточные и колониальные формы, у которых клетки кнаружи от цитоплазматической мембраны несут покров из окремнелых чешуек.
Род малломонас (Mallomonas), обычный в пресноводном планктоне, наиболее богат видами. Клетки различной формы покрыты окремнелыми чешуйками или чешуйками со щетинками. В качестве представителя рода можно рассмотреть один из наиболее крупных видов – Mallomonas eaudata, для которого подробно описана не только ультраструктура содержимого клетки, чешуек и щетинок, но и механизм их образования, освобождения и отложения на поверхность клетки. Такого рода исследований сравнительно немного.
М. caudata имеет два жгутика, из которых в оптическом микроскопе различим только один'. Этот жгутик нормального строения и несет два ряда волосовидных мастигонем. Второй жгутик, не различимый в световом микроскопе, выступает из клетки на короткое расстояние и скрыт покровом чешуек; он обладает аксонемой 9 – 9 + 0.
 | 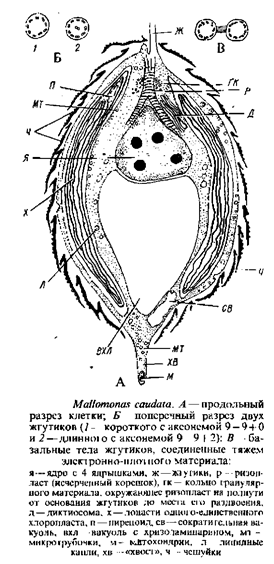 | ||
Два базальных тела соединены тяжем электронно-плотного материала. Ризопласт соединяет базальные тела с ядром, делясь непосредственно над ядром на две ветви. Несколько выше разветвления поперечно исчерченный корень окружен кольцом гранулярного материала, имеются и микротрубчатые корни. Одно ядро, содержащее до четырех ядрышек и расположенное по соседству с основаниями жгутиков, окружено оболочкой из двух мембран; наружная мембрана соединена с эндоплазматической сетью вокруг пластиды. Вблизи ядра – аппарат Гольджи. В клетке один двухлопастный хроматофор. Он окружен двойной мембраной и хлоропластной эндоплазматической сетью, содержит трехтилакоидные ламеллы. Имеется одна опоясывающая ламелла, в одной из лопастей хлоропласта находится пиреноид, который не удается наблюдать в световом микроскопе. Треть или половину заднего конца клетки занимает крупный пузырь с хризоламинарином. В цитоплазме содержатся еще жир, сократительные вакуоли, митохондрии, микротрубочки, слизевые тельца, которые располагаются непосредственно под цитоплазматической мембраной и состоят из темноокрашенной сердцевины, заключенной в мембрану. Предполагают, что они ответственны за слипание чешуек у М. eaudata.
Клетка покрыта кремнеземными чешуйками, расположенными по спирали. Каждая чешуйка отчасти окружена загнутым краем, отсутствующим в передней части, с многочисленными мелкими порами, почти в центре чешуйки – продолговатое отверстие. Щетинка имеет вид полой трубки с расщелиной на верхушке и несколькими зубцами по одному краю. Своим изогнутым основанием щетинка укреплена в дистальной части чешуйки, где отсутствуют поры. Место образования чешуек и щетинок – пространство между наружной поверхностью хлоропласта и плазмалеммой. Они формируются внутри плоских пузырьков – везикул, которые отпочковываются от хлоропластной эндоплазматической сети и сливаются с везикулами, являющимися дериватами аппарата Гольджи. Концы плоских везикул, производящих чешуйки, вздуты. Примордии и чешуек и щетинок представляют собой плоские пластины. На более поздних стадиях плоские пузыри с пластинами скручиваются в разной степени: в случае щетинок закручивание сильнее. Во время закручивания к везикулам прикрепляются трубчатые элементы, возможно принимающие участие в механизме закручивания. Следствием этого процесса является то, что в трубчатой части щетинки остается щель вдоль одной ее стороны. Клетка продуцирует или чешуйки, или щетинки, но не обе структуры одновременно. Мембрана везикула, содержащего зрелую чешуйку, сливается с плазмалеммой и чешуйка выталкивается в любом месте клеточной поверхности под существующим слоем чешуек. Возможно, чешуйки фиксируются в определенном положении слизью, которая вырабатывается и извергается наружу тельцами, расположенными непосредственно под плазмалеммой. Каким образом чешуйки выстраиваются в правильном порядке по поверхности клетки, пока не ясно. Освобождение щетинок происходит также посредством экзоцитоза. Первой выталкивается из клетки верхушка щетинки; у более длинных щетинок верхушка должна быть вытолкнута из клетки еще тогда, когда ее базальная часть находится в процессе формирования.