Страница
2

4. Структура пробки, всегда образующейся в результате активности аппарата Гольджи. у разных видов неодинакова. Пробка может быть составлена из разного количества слоев морфологически различающихся материалов, Каждый слой заключает несколько компонентов. Так, пробка у Uroglena и, возможно, у Mallomonas caudata содержит только первичный слой. У Ochromonas tubercuiata имеется периферический вторичный слой, состоящий из фиброзного материала, заключенного внутри везикул. У Dtnabryon cylindricum, D. divergens пробка цисты также включает второй периферический слой, но состоящий из гранулярного материала. У D. cylindricum имеется еще и третий слой пробки фиброзный. В химическом отношении пробка представляет собой полисахарид и не содержит вообще или содержит лишь небольшие количества кремнезема.
5. Цисты Ochromonas tubercuiata и Maltomonas caudata заключают по одному ядру и одной пластиде, как и вегетативные клетки. Цисты Dinobryon и Uroglena в отличие от вегетативных клеток содержат по две пластиды и по два ядра. Репликация этих органелл у изученных Сэндреном видов предшествует образованию цист, которые таким образом возникают бесполым путем. Это не исключает возможности появления цист у других видов в результате полового процесса. Такие случаи на уровне оптического микроскопа неоднократно описывались.
При прорастании цисты у Ochromonas tubercuiata пробка исчезает и содержимое выходит через пору.
Отдел золотистые водоросли (Chrysophyta) включает невколько классов, из которых ниже приводятся два: класс хризофициевые, или гетерохризофициевые, и сравнительно недавно установленный класс гаптофициевые, или примнезиофициевые, для золотистых водорослей с гаптонемой.
Класс хризофициевые – chrysophyceae
Класс объединяет водоросли с различными типами организации таллома, которые и кладутся в основу деления его на порядки: хризомонадальные, объединяющие мопадные формы, ризохризидальные, представленные золотистыми водорослями с ризоподиальной структурой, хризокапсаль-ные, включающие пальмеллоидные формы, хризосферальные – коккоидные, феотамниальные – нитчатые. Ниже приводятся только два порядка: хризомонадальные и хризокапсальные.
Порядок хризомонадальные нли хромулинальные – Chrysomonadales, Chromulinales
Это самый обширный порядок, включающий водоросли монадной структуры, одноклеточные и колониальные. Систематика хризомонад базируется на числе и строении жгутиков и особенно на характере клеточных покровов. Имеются одной двужгутиковые формы, и прежде считалось, что первые являются исходными, более примитивными. Однако с помощью электронного микроскопа обнаружено, что у кажущихся одножгутиковых форм имеется второй маленький боковой жгутик. Было высказано предположение, что двужгутиковые хризомонады с гетероконтными и гетероморфными жгутиками могли быть исходными, а одножгутиковые формы возникли в результате редукции более короткого жгута. Клеточные покровы хризомонад различны: имеются формы голые, одетые лишь плазмалеммой, у других клетки заключены в целлюлозные домики, у третьих поверх плазмалеммы находится покров из окремнелых чешуек.
Размножение осуществляется делением клеток, у некоторых описан половой процесс.
Хризомонады – преимущественно пресноводные формы, чаще всего приуроченные к чистым водам. Обычно они встречаются в холодное время года, ранней весной и поздней осенью, некоторые – в зимнее время подо льдом. Однако, как было показано для синуры, температура воды здесь имеет лишь косвенное значение. Решающим же фактором является химизм воды, меняющийся на протяжении года: в холодное время года из-за отсутствия другой растительности в воде содержится больше железа и азота. Большинство этих водорослей живут в планктоне, имеют специальные приспособления к планктонному образу жизни и некоторые вызывают «цветение» воды, окрашивая ее в бурый цвет.
Ниже будут рассмотрены представители трех семейств: охромонадовые, или хромулиновые, динобриевые и синуровые.
Семейство Охромонадовые – Ochromonadaceae. Сюда относятся голые формы, клетки которых покрыты только цитоплазматической мембраной с одним или двумя неодинаковыми жгутиками.
Род охромонас, виды которого обычны в планктоне и нейстопе пресных вод, реже солоноватоводных водоемов, представлен одиночными, окрашенными в золотистый цвет клетками с двумя гетероконтными и гетероморфными жгутиками. Клетка охромонас голая, снаружи одетая только цитоплазматической мембраной. Каплевидная форма ее поддерживается цитоскелетом из периферически расположенных микротрубочек. В центре клетки находится клеточное ядро, окруженное ядерной оболочкой из двух мембран. В расширении между мембранами ядерной оболочки заключены два пластинчатых хроматофора с типичной для всего отдела ультраструктурой. В задней части клетки находится крупная вакуоль с хризоламинарииом, в цитоплазме рассеяны митохондрии, в передней части клетки – аппарат Гольджи. От переднего конца клетки отходят два неодинаковых по длине и форме жгутика. Г. Бак проследил тонкое строение и происхождение мастигонем у Ochromonas danica. Эта водоросль удобна для изучения динамики развития мастигонем, так как ее клетки легко теряют жгутики, а затем образуют их заново. Таким образом можно изучить материал на разных стадиях регенерации жгутикового аппарата. Оказалось, что сами мастигонемы у охромонас сложнее, чем у фукуса и аскофиллума, и в их сборке и выведении на поверхность клетки участвует не только перинуклеарное пространство, но и элементы аппарата Гольджи. Готовые трубчатые мастигонемы охромонас состоят из базальной части, прикрепляющейся к мембране жгута, микротрубчатой части и 1–3 терминальных нитей. Кроме этих компонентов, сходных с таковыми мастигонем бурых водорослей, трубчатый стержень мастигонемы по всей поверхности несет длинные и короткие латеральные нити. Трубчатые мастигонемы располагаются на длинном жгутике в два неодинаковых ряда: с одной стороны мастигонемы прикрепляются к специальным участкам жгутиковой мембраны поодиночке, а на противоположной стороне жгута – пучками по три – пять. Помимо трубчатых мастигонем, длинный жгут несет тонкие простые нитевидные фиброзные мастигонемы. Короткий жгутик имеет только фиброзные мастигонемы. На рис. 16, В представлена схема возможной последовательности сборки мастигонем и их освобождения при регенерации жгутика у охромонас. По мере удлинения жгута новые мастигонемы и, возможно, мембрана, к которой они прикрепляются, добавляются к основанию жгута.
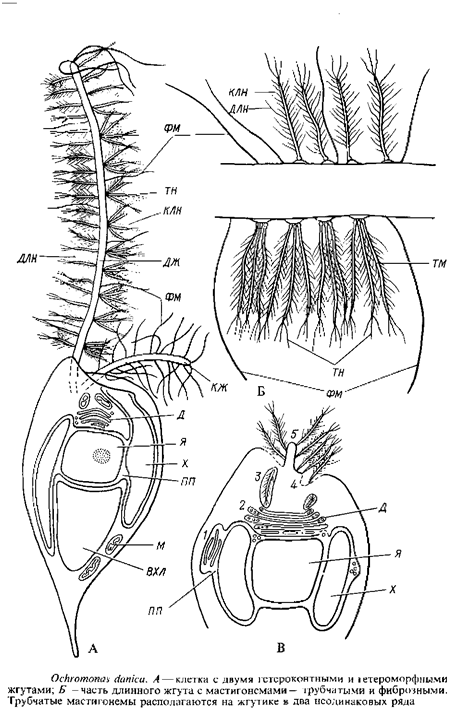
Такое же образование промастигонем и выделение готовых мастигонем поодиночке или в группах, наружу к основанию жгутика в цистернах аппарата Гольджи наблюдалось и у Ochromonas minuta. Жгутиковый корень у охромонаса состоит из поперечно исчерченных пучков фибрилл, которые, разветвляясь, достигают поверхности клеточного ядра.
Перед митозом базальные тела двух жгутиков реплицируются, хлоропласт делится перетяжкой, во время профазы реплицируются диктиосомы, появляется второй ризопласт. Вокруг ядра увеличивается число микротрубочек, ядерная оболочка начинает разрушаться у полюсов, что позволяет микротрубочкам войти внутрь ядра и образовать веретено. С каждым полюсом ядра ассоциируется по одному ризопласту, к которому, по-видимому, прикреплены трубочки веретена. Одни трубочки веретена простираются от полюса к полюсу, другие присоединяются к хромосомам. Остатки ядерной оболочки разрушаются, за исключением участков, соседних с хлоропластами. Хлоропластная эндоплазматическая сеть остается интактной на протяжении всего митоза. В анафазе микротрубочки удлиняются и хромосомы разъединяются на две группы. В телофазе появляются ядрышки и ядерные оболочки. Цитокинез происходит путем продольного раскола, который начинается у переднего конца клетки между двумя нарами жгутиковых оснований. Об образовании цист у Ochromonas сказано выше.