Обзор методов и способов измерения физико-механических параметров рыбы
Необходимое число экспериментов можно определить следующим образом. Сначала провести пять-шесть экспериментов и рассчитать доверительный интервал, а затем уточнить необходимое количество измерений, исходя из того, что уменьшение доверительного интервала в λ раз обеспечивает увеличение количества измерений в λ 2 раз.
В табл. 1 приведены результаты опытов по определению коэффициен тов трения покоя атлантической сельди по нержавеющей стали при продолжительности неподвижного контакта, равной 10 с. Все эксперименты по определению коэффициентов трения покоя проводились в зависимости от продолжительности непод вижного контакта. Время неподвижного контакта принималось равным 0, 10, 20, 30, 40, 60, 120, 180, 240-и 300 с. Как было установлено, изменение силы трения практически происходит в интервале от 0 до 300 с. Коэффициенты трения покоя зависят от массы рыбы и температуры ее тела. При проведении экспериментов брали рыб массой, наиболее характерной для данного вида. Поскольку коэффициент трения покоя изменяется обратно пропорционально корню кубическому из массы рыбы, то при изменении массы рыбы в 1,5 раза коэффициент трения изменяется в 1,13 раза. Все эксперименты проводили при температуре 10—12° С.
Кинетические коэффициенты трения рыб.
Кинетические коэффициенты вычисляются как частное от деления силы трения на силу нормального давления. Силы трения определяются в зависимости от скорости скольжения на экспериментальных установках, описанных в разделе «Установки для исследования коэффициентов трения». Силы трения определяются по осциллограмме для того момента, когда фрикционный контакт полностью сформирован.
Анализируя осциллограмму процесса трения, можно еще раз убедиться в том, что сила трения зависит от площади фрикционного контакта. В начальный момент сила трения меньше, чем в конце; по мере формирования площади фрикционного кон такта она увеличивается. Площадь фрикционного контакта при движении формируется значительно быстрее, чем при покое. Ее формирование практически заканчивается за 5—6 с. В интервале исследованных скоростей будет иметь место полужидкостное трение.
В процессе взаимного перемещения между рыбой и соответствующим материалом устанавливаются фрикционные связи, которые непрерывно возникают и разрушаются. Количество установившихся связей зависит от относительной скорости скольжения, формы тела рыбы (т. е. площади фрикционного кон такта) и свойств исследуемого материала. Как правило, с увеличением скорости сила трения, а, следовательно, коэффициент трения уменьшаются. Это объясняется тем, что с увеличением скорости относительного перемещения уменьшается количество связей между рыбой и исследуемым материалом, а, следовательно, и коэффициентом трения. По-видимому,при скорости более 2 м/с имеет место жидкостный режим трения, при котором с увеличением скорости воз растает сила трения.
ЭКСПЕРИМЕНТАЛЬНЫЕ ДАННЫЕ О КОЭФФИЦИЕНТАХ ТРЕНИЯ РЫБ
Коэффициенты трения единичных экземпляров рыб.
Были проведены многочисленные исследования по определению коэффициентов трения единичных экземпляров некоторых видов рыб по материалам, наиболее часто используемым для изготовления рабочих органов рыбообрабатывающих машин.
Экспериментальные исследования по определению коэффициентов трения и кинематических коэффициентов трения про водили на установках, описанных в разделе «Установки для исследования коэффициентов трения».
Опыты проводили только с размороженной рыбой. Для достоверности результатов опыты по определению коэффициентов трения при одних и тех же продолжительности неподвижного контакта и скорости повторяли 15—20 раз, а затем определяли средние значения и средние квадратичные отклонения. По сред ним значениям строили графические зависимости коэффициен тов трения покоя от продолжительности неподвижного кон такта,
На рис. 5 представлены графические зависимости коэффициентов трения покоя сельди по нержавеющей стали, оргстеклу и прорезиненной лейте от продолжительности неподвижного контакта. Как видно из приведенных данных, наименьший коэффициент трения покоя получен при контакте сельди с оргстеклом, несколько больший — при контакте с прорезиненной лентой. Оргстекло имеет чистоту поверхности, соответствующую седьмому, а нержавеющая сталь — пятому классу шероховатости. При контакте рыбы с нержавеющей сталью и оргстеклом сила трения, по-видимому, в основном обусловливается адгезионной составляющей, поэтому коэффициенты трения сравнительно невелики.
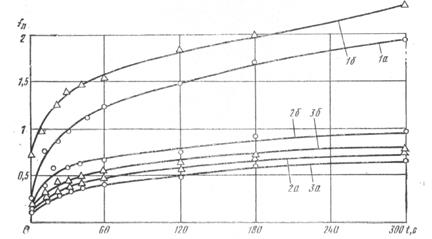
Рис. 5. Зависимость коэффициентов трения покоя сельди от продолжительности неподвижного контакта: 1 — сельдь — прорезиненная лента; 2 — сельдь — нержавеющая сталь; 3 — сельдь — оргстекло: а — при ориентации рыбы головой по направлению движения; б — то же, хвостом по направлению движения.
Прорезиненная лента является легко деформируемым материалом, при контакте с которым сила трения обусловливается деформационной и адгезионной составляющими, а поэтому и коэффициент трения для этого материалов значительно больше, чем для других. При ориентации сельди хвостом по направлению движения коэффициент трения будет несколько больше, чем при ее ориентации головой по направлению движения. Это объясняется строением чешуйчатого покрова. При ориентации рыбы хвостом по направлению движения увеличивается деформационная составляющая силы трения.
Коэффициенты трения рыб, замороженных в брикеты.
Для транспортировки рыбы, замороженной в брикеты, по наклонным плоскостям и посредством конвейеров необходимо знать углы наклона, при которых возможна такая транспортировка. Углы наклона можно определить, если известны коэффициен ты трения
tgφ = tgα = f (3)
где φ - угол трения; α - угол наклона плоскости; f - коэффициент трения.
Коэффициенты трения рыбы, замороженной в брикеты, определяли на тех же экспериментальных установках, что и коэффициенты трения покоя отдельных экземпляров рыб. Исследованные поверхности водой не смачивались. На рис. 6 и 7 представлены экспериментальные данные исследований коэффициентов трения рыб, замороженных в брикеты, по нержавеющей стали и прорезиненной ленте в зависимости от продолжительности неподвижного контакта. Анализ опытных данных позволяет предполагать, что между льдом и исследуемой поверхностью устанавливаются так называемые мостики сварки, которые и обусловливают величину силы трения.
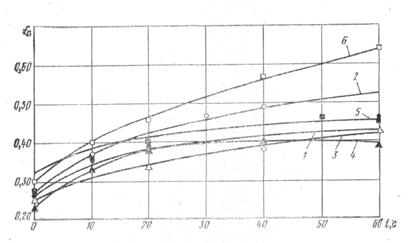
Рис. 6. Коэффициенты трения покоя различных видов рыб, замороженных в брикеты, в зависимости от продолжительности контакта с нержавеющей сталью:
1 — сельдь; 2 —треска; 3 —сайра; 4 — ставрида; 5 — скумбрия; 6 — сардинелла.
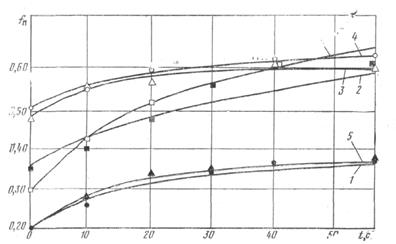
Рис.7. Коэффициенты трения покоя различных видов рыб, замороженных в брикеты, в зависимости от продолжительности неподвижного контакта с резиной: