Морфология колонии и распределительная система у двух видов герматипных кораллов рода acropora
На поверхности скелета ветви между его ребрами и столбовидными элементами находится подсистема поверхностных полостей, ограниченная с внешней стороны ценосарком (рис. 1, Б, Г, 8). На внешних краях элемента скелета происходит их нарастание (рис. 2, А), что приводит к формированию очередного “этажа” и замыканию полостей, лежавших первоначально на поверхности, т. е. приобщение их к подсистеме интеррадиальных полостей (рис.2, Б). Иными словами, каждая полость в интеррадиальной подсистеме была в свое время поверхностной.
Полости в колонии обычно выстланы живой тканью, что легко обнаружить по окрашиванию ее витальными красителями. Полости служат для прохода мезентериальных нитей (аконций), длина которых может приблизительно в 10 раз превышать длину кораллита. На сколах хорошо видно, как аконции движутся по проходам.
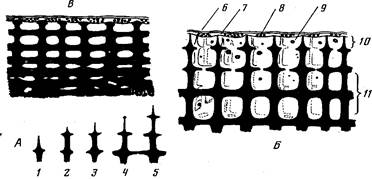
Рис. 2. Схема формирования и строения скелета на участке между кораллитами у поверхности ветви коралла рода Асrороrа, а также взаимное положение скелета и ценосарка (ткани, выстилающие скелет внутри колонии, не изображены): A—схема последовательных этапов роста столбовидных элементов скелета; Б—молодой участок ветви с острыми столбовидными элементами скелета; В—старый участок ветви с тупыми элементами скелета на поверхности и полностью заросшими полостями в центре ветви: /—5 — последовательные этапы роста элемента скелета; 6 — эктодерма ценосарка; 7— энтодерма ценосарка; 8 — зооксантсллы в энтодерме; 9 — мигрирующие клетки и частицы; 10—подсистема поверхностных полостей; П—подсистема интеррадиальных полостей
Очевидно, ткань, выстилающая скелет, не прекращает кальцификацию, так как со временем полости уменьшаются в размерах вплоть до полного их исчезновения (рис, 2, В). Скелет в старых частях колонии слабопористый, почти монолитный (рис. 1, Д). Естественно, что периферические полости в старой части колонии могут быть совсем молодыми и поэтому крупными. Из-за кальцификации центра ветви в старых ее частях транспорт веществ в них может осуществляться только по периферии, и главной в этом отношении становится подсистема поверхностных полостей.
Распределительная система в колонии
Система полостей в колонии определяет морфологическую основу распределительной системы. О распределительной системе у герматипных кораллов в литературе нет сведений. Эксперименты над акропорами позволили внести некоторую ясность по этому вопросу. С помощью инъекций мелких частиц активированного угля (использовались медицинские таблетки угля) удалось установить, что жидкость, заполняющая внутренние полости, находится в движении Сквозь покровы ценосарка и полипов было видно перемещение инъецированных частиц угля. По-видимому, движение внутренней жидкости осуществляется благодаря работе ресничек. Через ротовые отверстия полипов жидкость не выходит наружу колонии, т. е внутренняя среда достаточно замкнута.
Попытки косвенного изучения распределительной системы без повреждения ветви оказались безуспешными, поэтому была разработана простая методика для регистрации внутренних течений при вскрытии колонии. Часть ветви отсекали, переворачивали сколом кверху, и обе половинки укрепляли вертикально в аквариуме. Поверхности скола на обеих частях оказывались обращенными вверх. На каждую поверхность из пипетки наносили раствор витального красителя (нильский голубой). Краситель засасывался в те полости, через которые течение шло внутрь колонии, и окрашивал ткани, выстилающие эти полости. Напротив, там, где направление течения было из ветви, краска этим течением отгонялась, не приходила в соприкосновение с тканью и не окрашивала ее. Результаты эксперимента и его методика показаны на рис. 3.
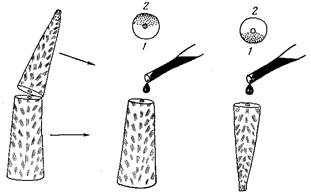
Рис. 3. Схема экспериментальной индикации постоянных течений внутри ветви у акропор с помощью витального красителя: 1,2 — изначально обращенные вверх (/) и к субстрату (2) стороны ветви на схеме поперечного скола. Точками отмечены окрасившиеся участки скола, в которых засасывающее течение было направлено внутрь ветви
Окраска показала, что на двух частях одной ветви окрашивались противоположно расположенные секторы. Это означает, что по одной стороне ветви течение идет к верхушке ветви, а по другой — к ее основанию. В аксиальном же канале преимущественно наблюдалось течение, направленное к основанию. Эксперимент, повторенный с толченым углем, дал такие же результаты. Непосредственное наблюдение под бинокуляром за исходящим течением по движению выносимых им частиц и клеток также подтвердило опыт. Эксперимент был расширен путем инъекции угля в проксимальную часть неповрежденной ветки. Благодаря тому, что в проксимальной части распределительная система ограничена периферическими полостями, здесь можно наблюдать за движением частиц угля под покровами достаточно долго, так как частицы не могут проникнуть в глубь колонии. Результат и этого опыта не противоречил полученным ранее данным. Следовательно, наблюдаемый эффект не является результатом повреждения ветви при ее расколе.
Описываемые опыты были проведены на колониях A. formosa. Затем эффект был проверен и на других видах: A. aspera, A cerealis (Dana, 1846), A. digiiifera, A. millepora (Ehrenberg, 1834), A, nasuia (Dana, 1846), A. pulchra, т. с. на видах с разным типом строения колонии. Везде результат был одинаков. Оказалось, что течение идет к верхушке по стороне ветви, обращенной вверх, а от верхушки к основанию — по стороне, обращенной к субстрату. Переворачивание ветви вокруг собственной оси на 180° не меняло хода течения. Следовательно, течение в распределительной системе определяется в процессе роста колонии и не зависит от положения ветви в эксперименте. Между секторами с дистально и проксимально направленными течениями были пограничные нейтральные зоны, в которых не были зарегистрированы течения.
Интересно, что данный тип распределительной системы — функционально билатеральный — никак не согласуется с радиальной симметрией системы полостей, служащей морфологической основой для распределительной системы. Следует в этой связи отметить, что течения внутриполостной жидкости на своем пути пересекают, не отклоняясь, все полости подряд, и полость кораллита оказывается такой же частью русла потока, как и все остальные полости.
Полученные результаты заставляют несколько изменить традиционное представление о колонии коралла—полипняке—как о продукте бесполого размножения. Колония у акропор, как это следует из приведенных данных, не агрегат зооидов, как-либо оставшихся в связи друг с другом из-за незавершенного почкования. Колония представляет собой безусловное целое с характерной формой, дифференциацией ма части и разделением функций.
Этот вывод может иметь значение и для систематики. Дифференциацию тела колонии на морфо-функциональные части можно использовать для более детального описания видов акропор, систематика которых до сих пор несовершенна. Давно известно (Яковлев, 1964; Wood-Jones, 1910), что герматипные кораллы, особенно акропоры, отличаются пластичной изменчивой формой, что затрудняет их систематику. Однако представление о вариабельности формы колонии во многом связано, во-первых, с отсутствием исследований морфологии колонии в целом, а во-вторых, с недостатком экспериментальных исследований зависимости формы колонии от генотипа и факторов внешней среды. Лишь после осуществления таких исследований можно будет делать выводы о степени пластичности формы колонии и о четкости границ между видами акропор.