Морфология колонии и распределительная система у двух видов герматипных кораллов рода acropora
У многих видов акропор рост радиальных кораллитов в проксимальных частях ветвей и особенно на их обращенных к субстрату сторонах настолько затормаживается, что скелет, формирующийся под ценосарком, начинает медленно обрастать кораллиты и в ряде случаев погребать их. Выглядит это как уменьшение длины кораллитов.
В период размножения в ветвях колонии происходит формирование половых продуктов, а затем, после оплодотворения, вынашнвание планул в гастральных полостях полипов. Верхушки же ветвей в размножении не участвуют, что служит дополнительным критерием подразделения ветви на самою ветвь и ее верхушку. В энтодерме зооидов и ценосарка ветви, особенно вблизи поверхности, находится множество зооксантелл.
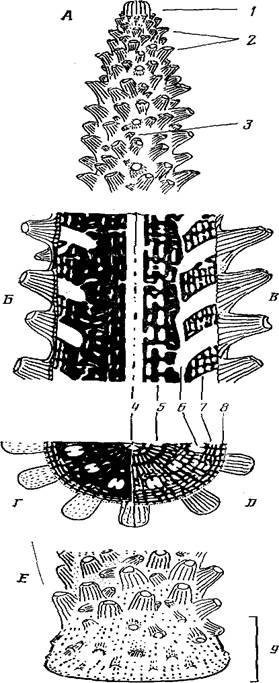
Рис. 1. Схема строения колонии Асrороrа formosa: A — дистальная часть ветви, включающая верхушку ветви; Б — Д — строение скелета и системы полостей на продольном (Б—В) и поперечном (Г— Д ) сечениях в дистальной и проксимальной (В, Д) частях; Е—основание колонии; / — аксиальный кораллит с формирующимися на боковых ребрах радиальными кораллитами 1-го порядка; 2—радиальные кораллиты 1-го порядка; 3—радиальные кораллиты 2-го порядка; 4 — аксиальный канал; 5 — подсистема циркумаксиальных полостей; 6 — полости радиальных кораллитов; 7— подсистема интеррадиальных полостей; 8 — подсистема поверхностных полостей; 9 — зона инкрустации
Ствол колонии. Ствол и ветви колонии морфологически и функционально похожи. Стволу колонии, как и ветвям, присущи: 1) кальцификация внутренних полостей в порядке от центра к периферии ствола; 2) наличие у ряда видов радиальных кораллитов 2-го порядка; 3) уменьшение размеров радиальных кораллитов вследствие их обрастания скелетом, формирующимся под ценосарком; 4) участие в половом размножении; 5) наличие массы зооксантелл в энтодерме. Отличительная особенность ствола — срастание между собой его ветвей, в результате чего образуется более мощная структура. Форма ствола из-за слияния с ветвями отлична от исходно цилиндрической. Скелет в местах слияний отчасти напоминает скелет в зоне инкрустации. Места срастания характеризуются повышенными кальцификацией и скоростью роста. Формируются как бы дополнительные зоны роста. Ствол колонии выполняет функцию опоры. Если ветви обычно легко отломить, то ствол сломать во много раз труднее, а без специальных инструментов и вовсе невозможно. По-видимому, в стволе происходит перекристаллизация скелета, которая приводит к усилению его прочности.
Основание колонии или зона инкрустации. По краю контакта ствола с субстратом имеется краевая зона или зона инкрустирующего роста (рис. 1, Е). Здесь осуществляется стелющийся рост колонии по субстрату. Формируется зона роста после метаморфоза планулы, следовательно, она свойственна колонии с момента се возникновения. В зоне инкрустации у многих видов акропор образуются радиальные кораллиты, ориентированные устьями к растущему краю. По ориентации радиальных кораллитов легко установить границу между стволом и основанием колонии (рис. I, E, 9).
Структура скелета радиальных кораллитов и остального скелета в зоне инкрустации обычно несколько отличается от таковой ствола. Подошву зоны инкрустации выстилает скелет, состоящий из особых элементов — шестигранных пластинок, плотно прилегающих друг к другу. Нигде более в нормальной колонии такие элементы не встречались. Благодаря разрастающемуся основанию колония прочнее прикрепляется к субстрату. Зона инкрустации, повторяя рельеф субстрата и проникая в мелкие полости, способствует усилению, прочности связи между колонией и субстратом. В зоне инкрустации возможно образование новых ветвей колонии на основе разрастания отдельных радиальных кораллитов. Для некоторых видов, например A. humilis (Dana, I846), это основной способ ветвления. Еще одна функция зоны инкрустации—предотвращение обрастания колонии другими кораллами. Конкурентная борьба между колониальными кораллами часто осуществляется при соприкосновении их оснований.
В ряде случаев инкрустирующий рост играет важную роль в процессе реколонизации. Нередко вся базальная часть колонии заносится песком и отмирает. После отступления песка выжившая часть колонии формирует зону инкрустирующего роста и обрастает затем свой же отмерший скелет. Так может повторяться много раз, и в строении скелета в соответствующей части колонии это находит ясное отражение. У разных видов Асrороrа соответствующие четыре части колонии развиты не в одинаковой пропорции. Так, A. formosa, A. pulchra (Brook, 1891), A. aspera имеют слабо выраженный ствол, но ярко выраженные ветви. Напротив, у A. hyacinthus (Dana, 1846) и A. clathrata (Brook, 1891) веточки маленькие, а ствол, представляющий продукт слияния ветвей, составляет основную часть тела колонии. У A. humilis и A. digitifera (Dana, 1846) ствол вообще сливается с зоной инкрустации и не может быть от нее отделен. Подобных примеров много, они требуют специального изучения. Для осуществления подобной работы необходим сбор целых колоний, а не нх фрагментов, при точном указании глубины, особенно вблизи поверхности рифа. Применение аквалангистской техники позволяет это осуществить.
Система полостей в скелете колонии
Многочисленные разноразмерные полости в скелете ветви Асrороrа расположены довольно упорядоченно. В размещении полостей прослеживается радиальная симметрия. Кроме того, полости расположены концентрическими окружностями (рис. 1, Г). Более подробное изучение системы полостей показало, что все они связаны между собой, но могут быть подразделены на пять основных подсистем.
В центре по оси ветви проходит аксиальный канал (рис. 1, А,Б,Г, 4). Полость его поделена шестью продольными септами на секторы. Аксиальный канал представляет продолжение гастралыюй полости аксиального кораллита, мезентерии которого простираются по каналу на расстояние нескольких сантиметров.
Два концентрических круга мелких полостей (рис. 1, А, Б, Г, 5) окружают аксиальный канал. Так как терминология для этих полостей не разработана, то можно называть нх подсистемой циркумаксиальных полостей. Скелет, окружающий аксиальный канал и эти полости, формируется в процессе роста аксиального кораллнта. Циркумаксиальные полости закладываются по периферии аксиального кораллита и являются таким же его продолжением, как и аксиальный канал.
Более крупные полости, выявленные на сколе то ближе, то дальше от оси ветви, — это полости радиальных кораллитов. В каждой такой полости можно обнаружить пару направляющих септ. Полости различаются на поперечном сколе по размерам: чем они ближе к центру, тем мельче. Это, во-первых, объясняется тем, что полости у радиальных кораллитов расширяются к устью и, следовательно, чем дальше от устья рассматривать сечение полости, тем оно мельче. Во-вторых, плоскость поперечного скола по-разному сечет полость изогнутого радиального кораллита: чем ближе к периферии, тем более касательно.
Полости радиальных кораллитов со всех сторон окружены подсистемой интеррадиальных полостей (рис. 1, Б, Г, 7). Эти полости делают скелет пористым, и благодаря им осуществляется связь между кораллитами. Скелет, в котором расположены интеррадиальные полости, состоит из радиалыю расходящихся стенок с отверстиями. Стенки довольно регулярно соединены между собой поперечными балками.