Метамерия или сегментация в живой природе
Метамерия может быть простой и сложной. О сложной метамерии говорят в тех случаях, когда составляющие тело животного метамеры располагаются повторяющимися группами, построенными по одному плану, так что эти группы являются как бы метамерами 2-го порядка.
Метамерия может возникать в организации животных несколькими путями: метамерным упорядочением первоначально хаотично расположенных гомотипных частей, (принадлежащих к одному и тому же типу частей); метаменрной дифференцировкой вдоль первоначально однородного целого; соединение первоначально независимых гомотипных частей. Первый и второй способ возникновения метамерии имеют между собой то общее, что здесь метамерное целое возникает из такого же целого либо вовсе лишенного метамерии, либо обладающего неопределённой поступательной симметрией. В третьем способе метамерное целое возникает из совокупности независимых между собой неметамерных единиц низшего порядка.
Метамерию мы встречаем во всём животном царстве. У простейших она наиболее ярко выражена у FORАMINIFERA. Здесь она возникает путём неполного деления и выражается в строении раковины, состоящей из ряда метамерно расположенных камер. Эта метамерия является несколько гетерономной, так как камеры не имеют одинакового размера. У CILIATA метамерия развивается путём стробиляции – возникновением метамерного целого путём незавершённого до конца поперечного деления. Деление особи происходит очень быстро и не успевает дойти до конца, как наступает следующее деление. Возникшие цепочки непрерывно распадаются, но животное всё равно сохраняет вид цепочки. Существует также метамерия колоний. У турбеллярий метамерия иногда возникает путём упорядочения – благодаря тому, что кишечник имеет разветвлённое строение половые железы, лежащие в свободных промежутках, приобретают столь же правильное расположение. Нервный аппарат в силу этого приобретает такое же метамерное строение.
У кольчатых червей (ANNELIDA) существует два способа роста: олигомерное (все сегменты полностью закладываются на стадии личинки, их число не велико и их дельнейшего образования не бывает) и полимерное (на протяжении оральной стороны закладывается лишь часть сегментов. В результате этих процессов создаётся вторая личиночная стадия – метатрохофора или нектохета. Вслед за тем у личинки образуется зона роста, в которой идёт усиленное деление клеток, ведущее к образованию нового отдела туловища). У олигомеров метамерия не распространяется на целомическую мезодерму, в то время как у полимеров она бывает метамерной. Существует три теории происхождения метамерии у аннелид: теория происхождения метамерии аннелид от антимерии кишечнополостных, теория возникновения метамерии путём метамерной дифференцировки и упорядочения органов, теория происхождения её путём подавленного поперечного деления.
В то время когда для кольчатых червей характерна, в основном, гомономная метамерия и гетерономность в пределах этого типа играет лишь подчинённую роль, членистоногие являются животными гетерономно-метамерными. В тоже время и самый характер гетерономности несколько различен, что в значительной мере зависит от различий в характере покровов и функциях конечностей. Наличие толстой кутикулы у членистоногих ведёт к образованию щитов, объединяющих группы смежных сегментов в отделы и слиянию сегментов между собой. Параподии многощетинковых выполняют локомоторную, гидрокинетическую, дыхательную и чувствительную функции. Примитивные конечности членистоногих несут, кроме того, хватательною и жевательную функции. Для членистоногих характерно постоянное использование конечностей для хватания и жевания. Примитивность метамерии трилобитов заключается, прежде всего, в чрезвычайной гомономности их конечностей. За исключением нитевидных антенн остальные конечности у трилобитов по одному общему типу. Они сравнительно мало различаются у разных видов и очень сходны на различных сегментах тела. Конечность трилобита состоит из предкоксы, коксы, шестичленного телоподита и многочленистого, несущего жаберные листочки, преэпиподита, прикрепляющегося к предкоксе. Кокса, а иногда и часть члеников телоподита вооружены жевательными и хватательными эндитами.
Marella - одна из самых
массовых форм в уникальном
 | 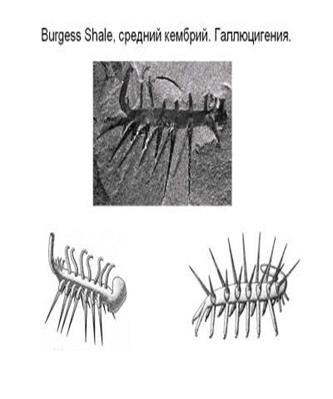 | ||
Ротовой диск Anomalocaris
Головные конечности отличаются от туловищных незначительно, чаще всего размерами телоподитов. Таким образом, по степени гомономности своих конечностей трилобиты не уступают бродячим полихетам.

Opabinia - существо неясного систематического положения
В направлении объединения сегментов тела в более крупные отделы – тагмы, трилобиты пошли несколько дальше. Максимальное число сегментов туловища у трилобитов – 42. У вышестоящих форм начинается дифференцировка туловища на две тагмы: грудь (торакс) и пигидий. Происходит этот процесс путём слияния тергитов задних сегментов в один пегидиальный щит, тогда как сегменты груди остаются свободными и подвижными. У многих трилобитов на поверхности пигидия ещё остаются поперечные борозды, указывающие на его метамерное строение, но у других они полностью исчезают, и пигидий представляется с дорсальной стороны совершенно однородным.
Вторую большую группу членистоногих представляют CHELICERATA. Для подтипа хелицеровых характерно слияние головных и грудных сегментов, в результате чего образуется единая головогрудь, состоящая из семи слившихся сегментов. Головогрудь несёт шесть пар конечностей, первая из которых видоизменилась в хелицеры, вторая пара конечностей видоизменилась в педипальпы. Остальные четыре пары являются ходильными ногами.
Их иногда разделяют на три класса: EURYPTEROIDEA, XIPHOOOSURA, ARACHNOIDEA. Все хелицеровые обладают высоко гетерономной
 |  |
 Ордовикский и силурийский ракоскорпионы, самые опасные морские хищники тех времен. Возможно, первые рыбы были "панцирными" для защиты от ракоскорпионов
Ордовикский и силурийский ракоскорпионы, самые опасные морские хищники тех времен. Возможно, первые рыбы были "панцирными" для защиты от ракоскорпионов