Математическое моделирование биосинтеза продуктов метаболизма
Если зависимость имеет возрастающий характер с насыщением, то зависимость удобно выразить в форме, похожей на уравнение Моно:
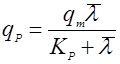 (25).
(25).
Если, наоборот, она падает с возрастом, то лучше подходит выражение, подобное уравнению Иерусалимского:
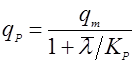 (26).
(26).
Если зависимость имеет экстремум, то оно может быть выражена, например, с помощью аппроксимирующего полиномиального уравнения [15]:
![]() (27).
(27).
Однозначная зависимость между qР и ![]() на практике встречается редко, часто зависимость скорости биосинтеза продукта от возраста учитывают в виде мультипликативного сомножителя, сопряженного с основной частью уравнения, учитывающего влияния остальных факторов.
на практике встречается редко, часто зависимость скорости биосинтеза продукта от возраста учитывают в виде мультипликативного сомножителя, сопряженного с основной частью уравнения, учитывающего влияния остальных факторов.
Модели деградации (инактивации) продуктов метаболизма.
Не всегда синтезированные продукты метаболизма остаются устойчивыми; часто они настолько нестабильны, что разрушаются уже в процессе самой ферментации. Поэтому, описывая материальный баланс по продукту метаболизма, необходимо учитывать кинетику его инактивации:
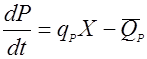 (28),
(28),
где ![]() – скорость деградации продукта метаболизма.
– скорость деградации продукта метаболизма.
При рассмотрении синтеза метаболитов, использовалась удельная скорость, в случае деградации, вводить удельную скорость не корректно, т.к. продукт существует отдельно от биомассы, и его деградация не зависит в общем случае от ее концентрации.
Рассмотрим модели кинетики деградации:
![]() (29),
(29),
деградация отсутствует.
![]() (30),
(30),
деградация идет с постоянной скоростью. Такое выражение странно выглядит в начале процесса, когда продукта еще нет; из уравнения же получается, что концентрация продукта снижается ниже нуля, что не имеет физического смысла.
![]() (31),
(31),
реакция разложения первого порядка, пропорционально количеству образовавшегося продукта [16].
![]() (32),
(32),
реакция разложения n-ого порядка, при чем n может быть как больше 1, так и меньше и не быть целым числом.
![]() (33),
(33),
реакция разложения зависит не только от концентрации продукта, но и от концентрации биомассы.
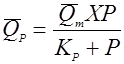 (34),
(34),
скорость реакции разложения зависит от концентрации биомассы и возрастает с концентрацией продукта до какого-то предела.
Приведенные уравнения инактивации (29)-(34) наиболее распространенные, существуют также и другие более сложные зависимости.
Модель накопления продукта метаболизма на примере лейцина.
L-лейцин- незаменимая аминокислота, необходимая для промышленного получения лизина. Производство лизина базируется на лейцинозависимых штаммах. Годовое производство лизина составляет приблизительно 500000 т/г. Лизин широко используется в с/х в качестве кормовой добавки. Лейцин также применяется в спортивном питании, т.к. является предшественником незаменимых жирных кислот, входящих в состав клеточных мембран.
Элементная формула L-лейцина (L-a-аминоизокапроновая кислота): C5H10NH2COOH.
Основным способом производства L-лейцина является микробиологический синтез с использованием штамма Corynebacterium glutamicum. Биосинтез проводился в лабораторном биореакторе. В отбираемых пробах определялась оптическая плотность – Х, содержание лейцина – P и содержание редуцирующих веществ (РВ) по Бертрану – S.
Полученные результаты приводятся в таблице 1.
Таблица 1.

Простейшим предположением о механизме микробиологического биосинтетического процесса является обобщение данных о том, что биосинтез, с одной стороны, ассоциирован с ростом, а с другой, осуществляется покоящейся клеткой.
Поэтому было решено использовать следующее соотношение (8):
 (35),
(35),
где P – концентрация продукта (лейцина), г/л;
aP, bP – эмпирические константы.
aP, bP были определены методом наименьших квадратов: aP=13.27, bP=1.249, сумма квадратов отклонений QP составила 0,884, а средняя квадратичная ошибка SP равна ± 0.6648.
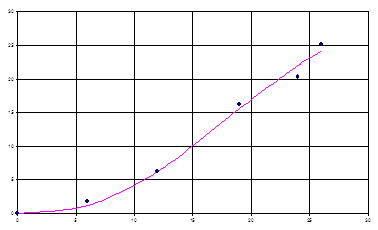 На рис. 3 показаны экспериментальные значения концентрации лейцина и модельные, из рисунка видно, что модель (8) в данном случае достаточно хорошо описывает биосинтез продуктов метаболизма.
На рис. 3 показаны экспериментальные значения концентрации лейцина и модельные, из рисунка видно, что модель (8) в данном случае достаточно хорошо описывает биосинтез продуктов метаболизма.
Рис. 2. Графическая интерпретация модели биосинтеза лейцина (35).
Список литературы
1. Арзамасцев А.А., Андреев А.А Математические модели кинетики микробного синтеза: возможности использования и новые подходы к разработке // Вестн. Тамб. ун-та. Серия: Естеств. и техн. науки.– 2000.– т.V., № 1– с. 111-130.
2. Renss M. Моделирование и оптимизация процессов // 8th Int. Biotechnol. Symp., Paris– 1988.– vol. 1.– p. 523-536.
3. Zeng An-Ping Кинетическая модель получения продуктов микробных клеток и клеток млекопитающих // Biotechnol. and Bioeng.– 1994.– vol. 45., N 4.– p. 314-324.
4. Vanrolleghem P.A. Структурный подход для выбора среди кандидатов в модель схемы метаболизма и установление неизвестных стехиометрических коэффициентов // Biotechnol. and Bioeng.– 1998.– vol. 2., N 3.– p. 133-138.
5. Перт С.Дж. Основы культивирования микроорганизмов и клеток.– М.: Мир.– 1978.
6. Luedeking R., Piret E.L. A kinetic study of the lactic acid fermentation: Batch process at controlled pH // J. Biochem. Microbiol. Technol. Eng.– 1959.– vol. 1., N 4.– p. 393-412.
7. Осипов Д.С., Гусельникова Т.В. и др. Математическая модель биосинтеза L-лейцина // Труды МГУИЭ.– 2001.– т.V.– с. 19-23.
8. Mori A., Terui G. Kinetic studies on submerged acetic acid fermentation: Inhibition by ethanol // J. Ferment. Technol.– 1972.– vol. 50, N 11.– p. 776-786.
9. Музыченко Л.А., Валуев В.И. Использование полунепрерывного культивирования микроорганизмов для получения продуктов биосинтеза // В кн.: Теория и практика непрерывного культивирования микроорганизмов.– Красноярск.– 1978.– с. 112-113.
10. Bajpai R.K., Reuss M. A mechanistic model for penicillin production // J. Chem. Technol. and Biotechnol.– 1980.– vol. 30,– p. 332-344.
11. Баснакьян И.А., Бирюков В.В., Крылов Ю.М. Математическое описание основных кинетических закономерностей процесса культивирования микроорганизмов // В кн.: Итоги науки и техники. Микробиология. Т. 5. Управляемое и непрерывное культивирование микроорганизмов.– М. – 1976.– с. 5-75.
12. Shu P. Mathematical model for product accumulation in microbiological processes // J. Biochem. Microbiol. Technol. Eng.– 1961.– vol. 3, N 1,– p. 95-109.