Источники и пути образования оксида азота в организме
Кроме того, для NO-синтаз характерна регуляция по механизму отрицательной обратной связи. При этом к действию оксида азота, выступающего в качестве неконкурентного ингибитора, более чувствительны конститутивные изоформы, снижение активности, которых происходит за счет связывания NO с атомом железа гемовой группы ферментов [11]. Ингибирующее действие NO на индуцибельную NOS, возможно, связано с ограничением димеризации молекулы фермента [3].
Считается, что NO может осуществлять ретроградную регуляцию не только путем взаимодействия с гемом фермента, но и через ингибирование транскрипции мРНК NO-синтазы (либо непосредственно, либо за счет угнетения активации фактора транскрипции NF-kB) [3] .
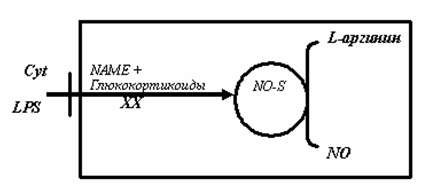
Рис.4. Стимуляция и ингибиция индуцибельной NO-синтазы [8].
Активация рецепторов гепатоцитов, клеток моноцитарно-макрофагальной системы и некоторых клеток крови цитокинами (Cyt) и липосахаридами (LPS), под влиянием которых происходит индукция индуцибельной синтазы окиси азота, синтезирующей NO из L-аргинина. Блокада (ХХ) i-NOS глюкокортикоидами или другими блокаторами ее активности. Образовавшийся NO диффундирует в межклеточное пространство [8],[9].
Таблица 2. Синтетические и природные агенты, модулирующие активность NO-синтаз [1].
|
Типы модулятора |
Изоформа фермента | ||
|
Noc-I |
Noc-II |
Noc-III | |
|
Активаторы/индукторы Ингибиторы |
Глутамат, N-метил-D-аспартат L-тиоцитруллин, 7-нитроиндазол, нитроаргинин |
Цис-платина, интерлейкин 1-β, интерферон-γ, липополисахарид, фактор некроза опухоли -α и –β, γ-излучение N-(3-(аминометил)-бензил)-ацетамидин, N6-(1-иминоэтил)-бензил)-ацетамидин; аминогуанидин, дексаметазон, ингибиторы сериновых протеаз, интерлейкин-4 и -10; L-NMMA; простагландин Е2, простациклин; N6-(1-иминоэтил)-L-лизин; 3-гидроксиантраниловая кислота; метотрексат |
Субстанция Р, АДФ, сдвиговое напряжение и пульсации давления крови, тромбин, гипомагнеземия, гистамин, Са2+-ионофоры, лейкотриены, олеиновая кислота, фактор активации тромбоцитов, аденозин, АТФ, ацетилхлин, брадикинин, серотонин; хлористый калий, эндотелин. Нитроимидоорнитин; иминоэтилорнитин, L-НАМЭ; АДМА |
Неферментативное образование оксида азота
Для биологических тканей помимо генерации оксида азота в ходе ферментативных реакций с участием NOS обнаружена возможность превращения нитрит-аниона в NO [7]. Этот процесс происходит в условиях ацидоза и при наличии восстановленных форм гемсодеращих белков, что характерно для такого патологического состояния как ишемия [3].
Так, образование оксида азота из нитрита может происходить в соответствии со следующей последовательностью реакций:
![]() NO-2 + H + HNO2
NO-2 + H + HNO2
![]() HNO2 {NOOH}
HNO2 {NOOH}
![]() {NOOH} + NO-2 N2O3 + OH-
{NOOH} + NO-2 N2O3 + OH-
![]() N2O3 NO + NO2
N2O3 NO + NO2
Кроме того, ионы NO-2 способны восстанавливаться до оксида азота в ходе окислительно-восстановительных реакций, акцептируя электроны с дезокси-форм гемсодержащих белков. Так, при взаимодействии NO с восстановленным гемоглобином происходит окисление Hb2+ до metHb и восстановление ионов NO-2 до NO:
![]() Hb2+ + NO-2 + 2H + metHb + NO + H2O
Hb2+ + NO-2 + 2H + metHb + NO + H2O
Нитритредуктазная активность также показана для миоглобина, цитохром-с-оксидазы и цитохрома Р-450.
Факт образования NO в биологических тканях из нитрит-аниона позволил предположить возможность существования механизма циклического превращения оксида азота в организме:
![]()
![]()
![]() L-Arg NO NO-2/NO-3 NO
L-Arg NO NO-2/NO-3 NO
Данное положение нашло отражение в концепции цикла оксида азота в организме млекопитающих. При этом NO-синтазная компонента обеспечивает эндогенный синтез NO, NO-2, NO-3 в присутствии кислорода. В условиях гипоксии или функциональной нагрузки, при которой осуществляется активное потребление кислорода, NO-синтазный механизм ингибируется [10].
В то же время дефицит кислорода приводит к активации нитритредуктазной компоненты цикла. Считается, что циклизация метаболических путей обеспечивает высокую степень упорядоченности и связанности систем биохимических реакций. Таким образом, механизм циклических превращений для NO и других высокореакционных азотсодержащих соединений гарантирует не только их эффективную переработку, но и достаточно быстрое выведение путем превращения в менее активные вещества, например ионы NO-2 и NO-3[3].
Методы определения оксида азота
Описанные в литературе методы определения NO можно условно разделить на прямые (таблица 2) и косвенные (таблица 3). В число первых входят те, с помощью которых осуществляется непосредственная регистрация NO, либо его комплексов. Прежде всего, это метод электронного парамагнитного резонанса (ЭПР) как средство изучения молекул с неспаренным электроном. Предложено использовать в качестве индикаторов NO регистрируемые методом ЭПР нитрозильные железосодержащие комплексы, устойчивые в биологически активных средах [1].
В живой ткани SH-содержащие белки, пептиды и аминокислоты образуют такие парамагнитные аддукты общего состава Fe(NO)2(SR)2, спектры ЭПР которых являются ассиметричными вариацией g-фактора от 2.01 до 2.05. однако из-за большого разнообразия естественных акцепторов NO и вариабельности их содержания, количественное определение этого радикала таким образом вряд ли возможно. В то же время гемопротеиды (гемоглобин, миоглобин, цитохром а3 и др.) образуют нитрозильные парамагнитные комплексы, имеющие широкий спектр ЭПР [1].
С разрешенной сверхтонкой структурой (СТС) в области значений g-фактора меньше 2. Анализ полученных спектров ЭПР свидетельствует о том, что структура указанных комплексов имеет ромбическую симметрию [13].
Таблица 3. прямые методы регистрации оксида азота[1].
|
Инструментальный метод |
Соединение-индикатор/реакция-индикатор |
Чувствительность |
|
ЭПР |
Fe-(NO)2-(SR)2 | |
|
ЭПР |
Hb-Fe(II)-NO |
5 мкМ |
|
ЭПР |
(ДТК)2-Fe(II)-NO |
1 мкМ |
|
ЭПР, L-линия |
Фьюзинит-NO |
1 мкМ |
|
Хемилюминесценция |
NO + O3 = hν + NOx |
20 нМ |
|
Амперовольтметрия |
NO + e = NOx |
10 нМ |