Страница
3
5. К моменту завершения синтеза первой цепи ДНК большая часть вирусной РНК разрушается РНКазой Н.
6. Синтез З'-конца второй цепи ДНК.
7. Удаление тРНК и оставшегося участка вирусной (+) РНК РНКазой Н.
8. Второй прыжок, в результате которого новосинтезированная вторая цепь ДНК комплементарно взаимодействует с тРНК-связывающей последовательностью первой цепи.
9. Удлинение З'-концов каждой цепи, образование дуплекса ДНК.
Вся последовательность реакций протекает без явного участия ферментов репликации клетки-хозяина (топоизомеразы, хеликазы, праймазы, ДНК-связывающего белка, лигазы и т.д.). При этом следует отметить, что молекулы вирусных ДНК длиннее молекул вирусных РНК, которые послужили матрицей для обратной транскрипции. Действительно, к 5'-концу (+) цепи вирусной ДНК добавилась последовательность U3, а к 3 -концу этой цепи — последовательность U5. В результате на концах молекулы вирус специфической ДНК появился длинный (несколько сотен нуклеотидов) концевой повтор (ДКП или LTR.), имеющий структуру U3U5.
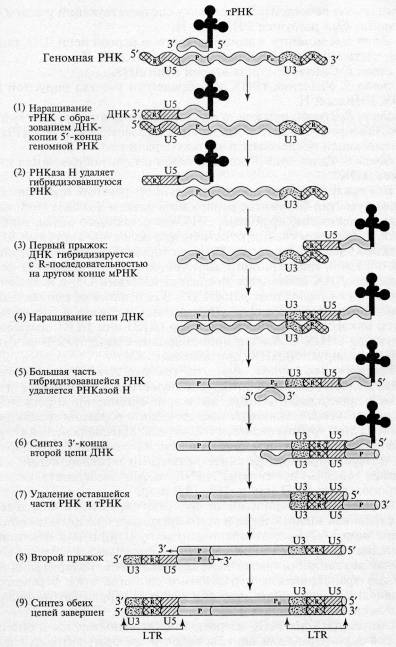
Рис.2. Схема обратной транскрипции ретровирусной РНК с образованием двуцепочечной ДНК (Darnell J., et.al. Molecular Cell Biology. – N. Y.: Scientific Amer. Books, 1986. – P. 1052)
Синтез ДНК на РНК-матрице in vitro и ревертаза используется в генетической инженерии для синтеза генов и их фрагментов, а также целенаправленного синтеза на матричных РНК комплементарных молекул ДНК (кДНК) для расшифровки первичной структуры РНК и белков.
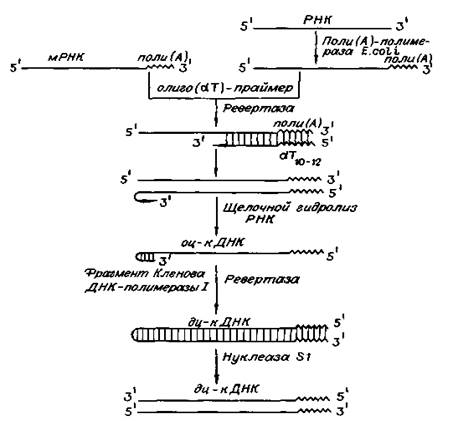
Рис. 3. Схема получения кДНК с использованием ревертазы вируса и трех дополнительных ферментов: поли (А)-полимеразы, фрагмента Кленова ДНК-полимеразы I и нуклеазы S1. (Цитировано по)
Реакцию обратной транскрипции проводят в специально подобранных условиях с использованием сильных ингибиторов РНКазной активности. При этом удается получать полноразмерные ДНК-копии целевых молекул РНК. В качестве праймера при обратной транскрипции поли (А) - содержащих мРНК используют олигo (dT)-праймер, а для молекул РНК, не имеющих З'-поли (А) концов, — химически синтезированные олигонуклеотиды, комплементарные З'-концу изучаемой РНК. После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (обычно применяют обработку щелочью) осуществляют синтез второй цепи ДНК. При этом используют способность ревертазы образовывать на 3'-концах одноцепочечных кДНК самокомплементарные шпильки, которые могут выполнять функции праймера. Матрицей служит первая цепь кДНК. Данная реакция может катализироваться как ревертазой, так и ДНК-полимеразой I E. coli. Показано, что сочетание этих двух ферментов позволяет повысить выход полноценных двухцепочечных молекул кДНК. По окончании синтеза первая и вторая цепи кДНК остаются ковалентно связанными петлей шпильки, служившей праймером при синтезе второй цепи. Эту петлю расщепляют эндонуклеазой S1, специфически разрушающей одноцепочечные участки нуклеиновых кислот. Образующиеся при этом концы не всегда оказываются тупыми, и для повышения эффективности последующего клонирования их репарируют до тупых с помощью фрагмента Кленова ДНК-полимеразы I E. сoli (остающаяся часть молекулы, которая сохраняет присущие ей каталитические активности).
Уже одно это открытие формально поставило РНК в центр основного постулата молекулярной генетики, так как показало, что поток генетической информации распространяется от РНК не в одном, а в двух направлениях: не только к белку, но и к ДНК. Все более глубокое проникновение в механизмы основных молекулярно-генетических процессов (репликацию, транскрипцию и трансляцию) способствовало возникновению понятия о неканонических функциях РНК, осознанию полифункциональности рибонуклеиновых кислот.
I.2 Функциональные возможности РНК
Для того чтобы оценить значение РНК в природе в целом, следует попытаться ответить на вопрос: что могут РНК?
К достаточно давно определенным каноническим функциям РНК относятся: способность выполнять роль мессенджера при передаче наследственной информации о структуре белка от ДНК к белоксинтезирующему аппарату клеток (мРНК), участвовать в формировании структуры рибосом (рРНК), обеспечивать специфическое акцептирование и перенос аминокислот к рибосомам (тРНК).
Вместе с тем РНК свойственны особые неканонические функции, реализуемые на разных этапах программы жизни тех или иных организмов.
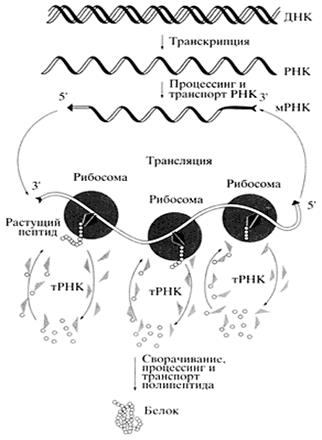
Рис. 4 Общая схема биосинтеза белка (Цитировано по Спирину А.С.)
В биосинтезе белка (трансляции) РНК безусловно играет определяющую роль. Различные по структуре рРНК формируют основу субчастиц рибосомы и определяют взаимодействие субчастиц при сборке полной рибосомы. Присоединение мРНК к рибосоме детерминируется комплементарным взаимодействием определенных участков мРНК и рРНК. Активация аминокислот, их специфическое акцептирование и доставка к рибосомам осуществляется тРНК. Кодон-антикодоновое взаимодействие между мРНК и тРНК обеспечивает перевод нуклеотидной последовательности информационных макромолекул в аминокислотную последовательность синтезируемых белков. Сама реакция образования пептидной связи (транс-пептидирование) и продвижение рибосомы по мРНК (транслокация) также, по всей видимости, связаны с функционированием рРНК. Пространственная структура мРНК непосредственно влияет на скорость трансляции, а ее способность взаимодействовать с разнообразными регуляторными белками, особенно характерная для высших эукариот, является основой для тонкой регуляции биосинтеза белка.
При «включении» гена происходит локальное расплетение спирали ДНК. Затем с гена, кодирующего белковую молекулу, синтезируется его РНК-копия. После ряда «превращений» она становится матричной РНК, т. е. матрицей для синтеза белка. мРНК переносится из ядра клетки в цитоплазму, где связывается с рибосомами, на которых и «производится» белок. Он синтезируется из активированных аминокислот, присоединенных к специальным транспортным РНК.
В процесс трансляции вовлечено множество макромолекул и макромолекулярных комплексов. При трансляции происходит считывание генетической информации, заключенной в мРНК, рибосомами и ее передача полипептидным цепям белков, т.е. биосинтез полипептидных цепей, последовательность аминокислот в которых определена последовательностью нуклеотидов в мРНК в соответствии с генетическим кодом.
Свободные аминокислоты не узнаются рибосомами. Чтобы это произошло, аминокислоты должны поступать в рибосомы в виде конъюгатов с тРНК (аминоацилированных тРНК), последовательности нуклеотидов которых распознаются аппаратом трансляции. В каждой молекуле тРНК имеется участок из трех нуклеотидов, комплементарный кодону мРНК. Именно эта последовательность, называемая антикодоном, определяет положение аминокислоты в полипептидной цепи. В ходе каждого индивидуального акта трансляции рибосома распознает кодон мРНК и в соответствии с ним выбирает аминоацилированную тРНК, антикодон которой соответствует транслируемому кодону. После этого происходит соединение посредством пептидной связи очередной аминокислоты с С-концевой аминокислотой растущей цепи полипептида.