Биологическая функция нуклеиновых кислот
Данная номенклатура нуклеотидов рассматривает их как фосфорные эфиры. В то же время благодаря наличию кислотной фосфатной группы удобно рассматривать нуклеозидмонофосфаты как кислотные производные исходных нуклеозидов, например, адениловая, уридиловая, гуанидиловая, цитидиловая кислоты.
Нуклеотиды — сильные кислоты, так как остаток фосфорной кислоты, входящей в их состав, сильно диссоциирован. При рН 7,0 свободные нуклеотиды в клетках находятся главным образом в форме
R —рибоза— ![]()
где R—азотистое основание.
Уникальны биохимические функции нуклеотидов. В качестве основных можно отметить следующие:
1) являются строительными блоками нуклеиновых кислот (ДНК и РНК); участвуют в молекулярных механизмах, с помощью
которых генетическая информация хранится, реплицируется и транскрибируется;
2) выполняют важную роль в энергетическом (фосфорном) обмене, в аккумулировании и переносе энергии;
3) служат агонами (коферментами и активными простетическими группами) в окислительно-восстановительных ферментах;
4) играют важную роль в синтезе олиго- и полисахаридов, жиров.
Таким образом, нуклеотиды — универсальные биомолекулы, играющие фундаментальную роль в обмене веществ и энергии живой клетки.
1.2. Первичная структура полинуклеотидов
ДНК и РНК представляют собой полинуклеотиды, имеющие три уровня структуры: первичную, вторичную, третичную.
Специфичность нуклеиновых кислот определяется не только их нуклеотидным составом, но и последовательностью отдельных нуклеотидов в цепи нуклеиновых кислот. В состав ДНК входит всего 4 нуклеотида, но, учитывая очень высокую молекулярную массу ДНК, нетрудно представить, что разнообразие ее типов выражается поистине астрономическими цифрами. Например, если мы возьмем цепочку, состоящую только из 100 нуклеотидов, то очевидно, что она может быть построена 4![]() способами.
способами.
Установлено, что ДНК каждого определенного вида характеризуется только ей присущей специфичной последовательностью нуклеотидов.

Рис.1 Схематическое изображение фрагмента полинуклеотида
Полинуклеотиды состоят из нуклеотидов, соединенных фосфорноэфирными связями с участием 3'- и 5'- углеродных атомов пентозных остатков двух соседних нуклеотидов. Длинные полинуклеотидные цепи содержат тысячи, миллионы нуклеотидных остатков. Фосфатные группы в цепях обладают сильнокислыми свойствами и при рН 7,0 полностью ионизированы. Поэтому в живых клетках нуклеиновые кислоты существуют в виде полианионов. Нуклеиновые кислоты плохо растворимы в растворах кислот. Они экстрагируются из разрушенных тканей и клеток растворами нейтральных солей или фенолом.
1.3. Вторичная и третичная структуры ДНК
Растворы ДНК характеризуются аномальной (структурной) вязкостью. В потоке обладают двойным лучепреломлением, что объясняется удлиненной формой молекул ДНК.
В расшифровку структуры ДНК большой вклад внесли исследования Э. Чаргаффа и его сотрудников (1945—1951 гг.). Для разделения оснований, полученных при кислотном гидролизе ДНК, Э. Чаргафф использовал метод хроматографии. Каждое из этих оснований было определено спектрофотометрически. Он впервые определенно заявил, что ДНК обладают выраженной видовой специфичностью. ДНК, выделенные из различных источников, отличаются друг от друга по соотношению входящих в их состав азотистых оснований. Э. Чаргафф сформулировал закономерности состава ДНК, известные под названием правил Чаргаффа. Независимо от происхождения ДНК эти закономерности представляются следующим образом:
1) количество молекул аденина равно количеству молекул тимина (А = Т);
2) количество молекул гуанина равно количеству молекул цитозина (G = С);
3) количество молекул пуриновых оснований равно количеству молекул пиримидиновых оснований (А + G = Т + С);
4) количество оснований с 6-аминогруппами в цепях ДНК равно количеству оснований с 6-гидроксигруппами (А + С = G + Т);
5) отношение (G+С)/(А+Т) резко отличается для разных видов ДНК, но постоянно для клетки одного вида; это отношение называется фактором специфичности.
Фактор специфичности одинаков для ДНК различных органов и тканей одного организма и практически не отличается у разных видов животных и растений в пределах одного класса. У высших растений и животных его величина находится в пределах 0,55—0,93; у бактерий — 0,35—2,73.
Правила Чаргаффа сыграли решающую роль в разработке проблем молекулярной биологии. Именно они легли в основу открытия строения ДНК, ее вторичной структуры.
Прежде чем приступить к рассмотрению этой структуры, необходимо отметить следующее. Интерес к проблеме изучения структуры ДНК возрос в связи с полной неясностью механизма воспроизведения (репликации) ДНК, который отличается очень высокой степенью точности. На основании уже полученных экспериментальных данных предполагалось, что генетическая информация в живой клетке зашифрована (т.е. записана с помощью определенного кода) в специфической последовательности оснований ДНК. Однако было неясно, как воспроизводится такая последовательность.
В 1953 г. Дж. Уотсон и Ф. Крик предложили модель структуры ДНК (рис.2). Она учитывала рентгеноструктурные данные Р. Франклин и М. Уилкинса и "эквимолярность" оснований, открытую Э. Чаргаффом. Модель Уотсона — Крика не только объяснила физико-химические свойства ДНК, но и дала основание высказать предположение о возможном механизме репликации ДНК. Согласно их выводу, молекула ДНК должна быть двухцепочечной. Каждое основание одной цепи "спарено" с лежащим в той же плоскости основанием второй полинуклеотидной цепи. Это спаривание специфично; поскольку количество оснований с гидроксигруппами равно количеству оснований с аминогруппами, то, по Уотсону и Крику, только определенные пары оснований входят в структуру так, что могут образовывать друг с другом водородные связи. Так как аденин содержит аминогруппу, а тимин — гидроксигруппу, цитозин и гуанин — соответственно эти же группы и поскольку они находятся в ДНК в эквимолекулярных количествах, то разрешенными являются только пары А-Т и G-С
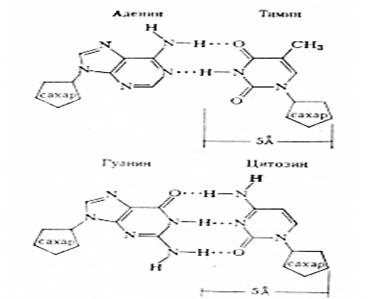
Рис.2. Соединение пар аденина и тимина, цитозина и гуанина в молекуле ДНК при помощи водородных связей
Между А и Т образуются две водородные связи, между G и С - три водородные связи.
Две полинуклеотидные цепи ДНК отличаются одна от другой как последовательностью оснований, так и нуклеотидным составом. Однако основания, стоящие в данном положении в одной цепи, определяют природу основания в другой цепи. Например, если в одной цепи стоит аденин, то напротив него в другой цепи будет располагаться тимин, и наоборот. Если в одной цепи стоит гуанин, то в другой обязательно будет цитозин, и наоборот. Действительно, рентгеноструктурный анализ ДНК показал, что пуриновые и пиримидиновые основания нуклеотидных остатков ДНК лежат в одной плоскости, перпендикулярной продольной оси молекулы, тогда как циклы дезоксирибозы находятся в плоскости, почти перпендикулярной той, в которой лежат циклы оснований.