Страница
3
Самой совершенной заменой зрения является эхолокация, при которой животное испускает высокочастотные сигналы и обнаруживает предметы по возникающему от них эху.

Рис.3. Строение лицевой части головы совы; некоторые перья удалены, чтобы показать асимметричное расположение ушей.
Принцип здесь такой же, как в военных радиолокаторах. Простые формы эхолокации встречаются у землероек, жирных козодоев и гималайских саланганов, которые ночуют и гнездятся в пещерах. Более совершенными ее формами обладают дельфины и другие морские млекопитающие, но своей вершины она достигает у летучих мышей.
После работы Гриффина появилось множество исследований механизма эхолокации у рукокрылых. Теперь мы неплохо знаем физиологию их издающего и принимающего сигналы аппаратов и механизмы мозга, участвующие в эхолокации. Читатель может найти более подробное их описание у Эверта и Гатри.
В процессе эхолокации летучие мыши испускают залпы ультразвуковых, т.е. не слышных для человека, импульсов малой длительности и высокой частоты. Такие короткие импульсы позволяют точно определять время образования эха, а значит, и расстояние до образующего его предмета.
Звуки, производимые другими животными и ветром, обычно бывают низкочастотными, поэтому маловероятно, чтобы на ультразвуковые сигналы летучих мышей накладывались помехи. Лабораторные опыты показали, что искусственные звуки частотой выше 20 кГц дезориентируют полет летучих мышей. Другое преимущество высоких частот состоит в возможности точной фокусировки, что делает возможным распознавание мелких объектов. Рукокрылые производят ультразвуковые сигналы особо устроенной гортанью и испускают их губами, как голоспинные листоносы Pteronotus, или из специальной формы ноздрей, как подковоносы Rhinolophus и представители семейства Phyllostomidae.
У летучих мышей много также специальных приспособлений, позволяющих им определять время и локализовать место возникновения эха от их ультразвуковых сигналов. У большинства рукокрылых, хватающих насекомых на лету, большие наружные уши, форма которых повышает дирекциональную чувствительность.
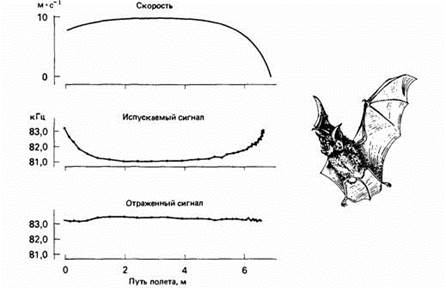
Рис.4. Эхолокация подковоноса, приближающегося к неподвижному предмету.
По мере приближения к нему скорость полета снижается. Летучая мышь приспосабливает частоту своих сигналов так, чтобы частота отражаемого ультразвука оставалась постоянной. После каждого испускаемого животным сильного сигнала эта чувствительность снижается специальными мышцами во внутреннем ухе. При очень коротких испускаемых импульсах, как у представителей сем. Vespertilionidae, конец импульса не перекрывается с началом его эха. Поскольку эхо приходит быстрее от близких объектов, импульсы постепенно укорачиваются по мере приближения объекта, и в результате такое перекрывание устраняется.
У других летучих мышей издаваемые импульсы и эхо от них перекрываются, поэтому, чтобы улучшить обнаружение эха, им нужны другие средства. Например, большой подковонос приспосабливает частоту своих сигналов так, чтобы частота возвращающегося эха поддерживалась в узких пределах. Для мыши, летящей к объекту, воспринимаемая частота эха всегда выше, чем частота испускаемого ультразвука. Это объясняется эффектом Допплера, возникающим вследствие относительного движения животного и объекта: чем быстрее они движутся навстречу друг другу, тем выше воспринимаемая частота излучения. Для компенсации такого эффекта летучие мыши меняют частоту своих сигналов так, чтобы воспринимаемая частота была как можно ближе к постоянной. Таким способом мышь может оценить скорость своего полета, а также направление и относительную скорость полета жертвы.
Как замена зрения эхолокационные способности рукокрылых производят большое впечатление. Лабораторные исследования показывают, что летучая мышь с размахом крыльев в 40 см может пролететь в полной темноте сквозь сетку с ячейками 14 на 14 см из нейлоновых нитей толщиной всего 80 мкм. Летучие мыши научались также ловить мелкие частички пищи, брошенные в воздух в полной темноте, и отличать съедобные предметы от несъедобных по небольшим особенностям их формы. Малая бурая ночница (Myotis lucifugus) может ловить на лету очень мелких насекомых, например плодовых мушек и комаров, с "результативностью" два насекомых в секунду.

Рис.5. Подковонос пролетает через сетку из нейлоновых нитей толщиной 80 мкм с ячейками 14 х 14 см. Слева - длительность и частота эхолокационных сигналов.
3. Зрительное опознание жертвы и хищника
Большинство хищников встречается с множеством разных видов, которые служат им добычей и которых необходимо отличать от других организмов. При этом чаще всего используются три признака - величина, движение и форма.
Когда хищники должны выбирать между особями-жертвами, отличающимися только размерами тела, они обычно хватают самую крупную. Эта стратегия наиболее эффективна в отношении затрат энергии. Однако по мере увеличения размера обычно наступает предел, за которым стимул больше не воспринимается как добыча. Например, когда обыкновенной жабе предъявляют как "добычу" объекты разной величины, она реагирует положительно на экземпляры, размеры которых находятся в определенных пределах, но активно избегает более крупные стимулы.
Как жаба судит о величине объекта? Проще всего делать это по величине изображения на сетчатке, измеряемого градусами угла зрения. Для объекта постоянной величины угол зрения меняется с расстоянием объекта от глаза. Близкие объекты выглядят крупнее, чем удаленные.
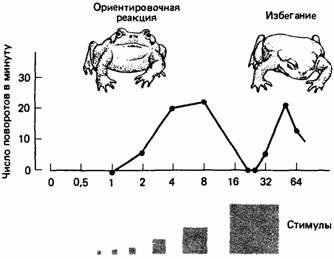
Рис.6. Жабы реагируют на маленькие движущиеся квадраты, ориентируясь в их направлении, но при определенных размерах квадрата эта реакция исчезает и большие квадраты вызывают реакцию избегания.
Чтобы выбрать добычу определенной величины, жабе нужно оценить абсолютную величину видимого объекта, учитывая и его размеры на сетчатке, и расстояние до него. Жабы-повитухи (Alytes obstetricans] в процессе развития постепенно научаются выбирать добычу постоянного размера. Сразу после метаморфоза они предпочитают искусственную приманку некоторого углового размера почти независимо от расстояния. Через шесть месяцев они ориентируются на объекты определенного абсолютного размера независимо от расстояния, т.е. за это время они как-то научаются включать суждения о расстоянии в свои оценки размеров. В некоторых случаях наблюдается склонность к более крупной добыче, но не из-за активного предпочтения, а потому, что она лучше различима. Так, радужные форели чаще ловят крупных ракообразных, чем мелких, потому что крупные видны на большем расстоянии.
Для того чтобы узнать добычу, некоторым видам, например лягушкам и жабам, нужно, чтобы она двигалась. Обыкновенная каракатица в норме нападает только на движущихся креветок. Но если у каракатицы отнять только что пойманную и парализованную ею креветку, то это головоногое немедленно снова нападет на нее даже на неподвижную. Некоторые хищники предпочитают добычу, которая движется беспорядочно. Так, личинки стрекоз предпочитают животных, совершающих зигзагообразные движения, а солнечная рыба скорее нападет на искусственную рыбку, если та извивается, чем на плавно движущийся объект. Иногда имеет значение соотношение формы и движения. Так, когда жабам предъявляют темную движущуюся полосу на белом фоне, они сразу же нападают на нее, когда полоса перемещается вдоль своей оси подобно червяку.