Эйкозаноиды - общая группа физиологически и фармакологически активных соединений
Простагландины PG можно рассматривать как производные гипотетической С2о-кислоты, получившей тривиальное название простаноевой:
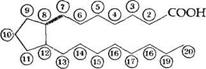
Простагландины обозначают символами, например PG А, где PG обозначает слово "простагландин", а буква А обозначает заместитель в пятичленном кольце в молекуле эйкозаноида.
Обнаружено шесть первичных природных простагландинов, три из них серии Е (PGE1, PGE2, PGE3) и три - серии F (PGF1a, PGF2a, PGF3a). Простагландины серии Е содержат в положении 9 кетогруппу, а простагландины серии F - гидроксигруппу. (Рис.2)
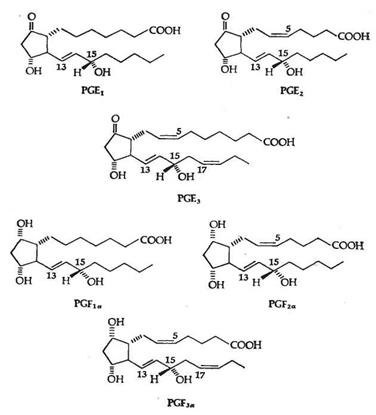
Рис.2 Шесть главных природных простагландинов
В дополнение к вышеприведенным шести первичным простагландинам имеются несколько вторичных простагландинов, большинство из которых являются продуктами ферментативного или химического превращения PGE. Характерным свойством простагландинов серии Е является их способность к дегидратации в PGA в кислой среде; образовавшиеся PGA затем в присутствии оснований изомеризуются в PGB, имеющие максимум поглощения при длине волны 237 нм. Эта последовательность реакций превращения PGE в PGB часто используется для идентификации PGE.
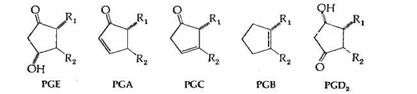
Простагландины серии F не претерпевают подобного превращения в кислой среде.
В зависимости от исходной жирной кислоты все эйкозаноиды делят на три группы:
1) Первая группа образуется из эйкозотриеновой кислоты. Хотя в пищевых продуктах этой кислоты нет, она способна образовываться в клетках при удлинении линолевой кислоты (С18: 3), которой много в растительных маслах. Для этой группы в соответствии с числом двойных связей простагландинам и тромбоксанам присваивается индекс 1: например, PgE1, PgI1
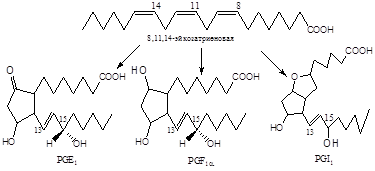
Вторая группа синтезируется из арахидоновой кислоты (С20: 4), по тому же правилу эйкозаноидам этой группы присваивается индекс 2. PgE2, PgI2.

Третья группа эйкозаноидов происходит из тимнодоновой кислоты (С20: 5). Эйкозаноидам этой группы присваивается индекс 3. Например, PgE3, PgI3
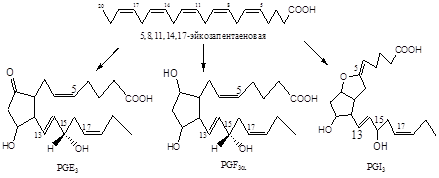
PG I - простациклины. Имеют 2 кольца в своей структуре: одно пятичленное, как и другие простагландины, а другое - с участием атома кислорода. Их также подразделяют в зависимости от количества двойных связей в радикалах (PG I2, PG I3).
Тромбоксаны. В отличие от простагландинов, тромбоксаны синтезируются только в тромбоцитах, откуда и происходит их название, и стимулируют их агрегацию при образовании тромба.
Тромбоксаны имеют шестичленное кольцо, включающее атом кислорода. Так же, как и другие эйкозаноиды, тромбоксаны могут содержать различное число двойных связей в боковых цепях и отличающиеся по активности. ТХ В2 - продукт катаболизма ТХ А2 и активностью не обладает.
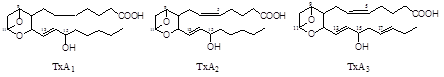
Рис.3. Структура тромбоксанов.
Биосинтез
После отделения арахидоновой кислоты от фосфолипида она выходит в цитозоль и в различных типах клеток превращается в разные эйкозаноиды. В клетках имеется 2 основных пути превращения арахидоновой кислоты: циклооксигеназный, приводящий к синтезу простагландинов, простациклинов и тромбоксанов, и липоксигеназный, заканчивающийся образованием лейкотриенов. (Рис.4)
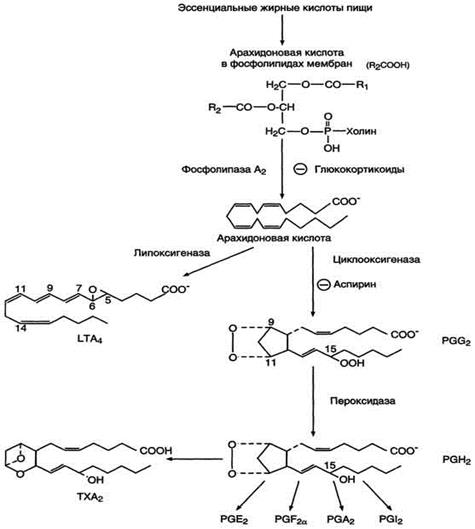
Рис.4 Синтез эйкозаноидов из арахидоновои кислоты.
Циклооксигеназный путь: синтез простагландинов и тромбоксанов
С20-три-, тетра - и пентаеновая жирные кислоты, служащие предшественниками простагландинов, входят в состав внутриклеточных фосфоглицеридов, из которых они высвобождаются под действием фосфолипазы А2. Активация фосфолипаз, ассоциированных с мембранами, происходит под действием многих факторов: гормонов, гистамина, цитокинов, механического воздействия. Этот процесс высвобождения предшественника, вероятно, играет роль регулирующей стадии в биосинтезе простагландинов, с помощью которой варьирует количество субстрата, подвергающегося последующему действию простагландинсинтазы. Последняя представляет собой связанный в мембране мультиферментный комплекс, содержащийся в микросомальной фракции тканевых гомогенатов. Необходимо отметить, что ингибиторное действие кортикостероидов на биосинтез простагландинов, связанное с торможением активности фосфолипазы А2, может служить объяснением антивоспалительного действия этих стероидных гормонов.
Фермент, катализирующий первый этап синтеза простагландинов, называется PG Н2 синтазой и имеет 2 каталитических центра. Один из них называют циклооксигеназой, другой - пероксидазой. Этот фермент представляет собой димер гликопротеинов, состоящий из идентичных полипептидных цепей. Фермент имеет гидрофобный домен, погружённый в липидный слой мембран ЭР, и каталитический домен, обращённый в полость ЭР. В активном центре циклооксигеназы находится тирозин (385), в активном центре пероксидазы - простетическая группа - гем. В организме имеются 2 типа циклооксигеназ (PGН2 синтаз). Циклооксигеназа 1 - конститутивный фермент, синтезирующийся с постоянной скоростью. Синтез циклооксигеназы 2 увеличивается при воспалении и индуцируется соответствующими медиаторами - цитокинами. Оба типа циклооксигеназ катализируют включение 4 атомов кислорода в арахидоновую кислоту и формирование пятичленного кольца. В результате образуется нестабильное гидропероксидпроизводное, называемое PG G2. Гидропероксид у 15-го атома углерода быстро восстанавливается до гидроксильной группы пероксидазой с образованием PG Н2. До образования PG Н2 путь синтеза разных типов простагландинов одинаков. Дальнейшие превращения PG Н2 специфичны для каждого типа клеток.
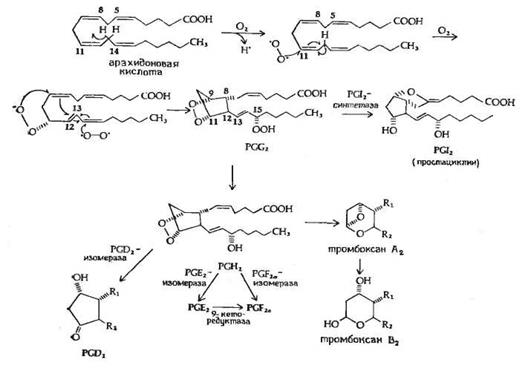
Рис.5. Предполагаемый механизм биосинтеза PGE2 и PGF2a из арахидоновой кислоты.
Биосинтез PGE2 и PGF2a, представленный на рис.5, начинается с арахидоновой кислоты и катализируется циклооксигеназой (М 70 000) жирных кислот, при этом образуется 15-оксиперокси-9-11-эндопероксид PGG2. Первая стадия в этой последовательности реакций приводит к потере атома водорода у С-13 с образованием свободного радикала. Эта стадия лимитирует скорость всего процесса. Затем происходят присоединение пероксидного радикала в положения 9 и 11 и образование 8,12 углерод-углеродной связи, которому предшествует изомеризация 11,12-двойной связи в 12,13-двойную связь и присоединение другого пероксидного радикала в положение 15, приводящее к изомеризации 12,13-двойной связи в 13,14-двойную связь. Все эти превращения рассматриваются как процесс одноэлектронного переноса, обозначенный на рисунке изогнутыми стрелками. Первый продукт этой последовательности реакций - это 15-оксиперокси-9,11-эндопероксид PGG2, существование которого было постулировано за несколько лет до его выделения и идентификации. PGG2 затем превращается в PGI2 или PGH2, вероятно, с помощью реакции пероксидазного типа. PGH2 является предшественником PGE2 и PGF2a в семенных пузырьках и других органах, а также тромбоксана А2 в тромбоцитах и легких.