Что и как закодировано в мРНК
Яркий пример регуляции трансляции мРНК в эукариотических клетках получен при изучении механизмов поддержания в клетках уровня свободного железа. Железо входит в состав активных центров очень многих белков, таких, например, как гемоглобин , миоглобин, цитохромы , однако ионы свободного железа токсичны для клетки и поэтому связываются и переводятся в нетоксичную форму белком ферритином. Синтез ферритина в клетке, в свою очередь, зависит от уровня свободного железа: в присутствии железа ферритин синтезируется, в то время как при его недостатке трансляция мРНК ферритина останавливается на стадии инициации. Выяснилось, что регуляция синтеза ферритина целиком зависит от специфической последовательности длиной 26 нуклеотидов, образующей шпилечную структуру в 5'-НТО мРНК ферритина (рис. 4, а). Этот регуляторный элемент при отсутствии железа связывается со специфическим белком, который препятствует сканированию 5'-НТО рибосомами и, таким образом, подавляет трансляцию мРНК на стадии ее инициации. Этот белок- репрессор имеет сродство к ионам железа и при связывании с ними перестает связываться с ферритиновой мРНК. После диссоциации белка мРНК становится активной в синтезе ферритина.
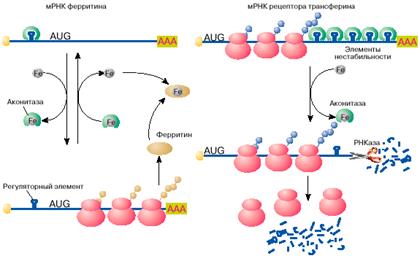
Рис. 4. Регуляция железом трансляции мРНК ферритина и стабильности мРНК рецептора трансферина
Вновь синтезированный ферритин отнимает железо у репрессора. Утратив железо, репрессор опять приобретает сродство к регуляторному элементу ферритиновой мРНК, связывается с ним и останавливает синтез ферритина. Большим сюрпризом явилось то, что белок-репрессор оказался хорошо известным ферментом цикла Кребса - аконитазой.
ОСОБЫЕ СТРУКТУРНЫЕ ЭЛЕМЕНТЫ мРНК ОПРЕДЕЛЯЮТ ВРЕМЯ ЖИЗНИ мРНК В КЛЕТКЕ
Скорость синтеза белка зависит от количества соответствующей мРНК в клетке. Содержание мРНК, в свою очередь, определяется скоростью ее синтеза и распада. Механизмы регуляции транскрипции мРНК начали исследовать еще в начале 60-х годов, механизмы регуляции процесса ее деградации - значительно позже, всего 5-10 лет назад.
Время жизни разных мРНК в клетках эукариот варьирует в очень широких пределах. Период их полураспада составляет от десятков минут до десятка суток. Наиболее стабильными являются мРНК, на которых синтезируются белки, обеспечивающие функционирование организма как целого (например, мРНК сывороточного альбумина ). Некоторые мРНК распадаются на определенных стадиях клеточного цикла или определенных этапах процесса клеточной дифференцировки. Время жизни мРНК в клетке, равно как и момент их деградации, запрограммировано специфическими последовательностями или структурами в мРНК, часто в их 3'-нетранслируемой области. Эти последовательности (структуры) обычно узнаются специфическими белками. Их связывание с соответствующими белками обычно приводит к стабилизации мРНК.
Например, за деградацию гистоновых мРНК на определенной стадии клеточного цикла отвечает особая шпилечная структура в 3'-НТО - элемент нестабильности мРНК. Введение такой структуры в 3'-НТО очень стабильной глобиновой мРНК приводит к ее деградации одновременно с гистоновыми мРНК. Для того чтобы эта структура выполняла свою функцию элемента нестабильности, она должна находиться на определенном, достаточно близком расстоянии от терминирующего кодона. Дестабилизирующим действием в мРНК онкобелка c- myc обладает AU-богатая последовательность длиной 60-80 нуклеотидов в 3'-НТО этой мРНК. В этой последовательности был выявлен многократно повторяющийся мотив AUUUA, который и является элементом нестабильности. Сходная последовательность была обнаружена в 3'-НТО мРНК для другого онкобелка - c-fos. Однако в этой мРНК имеется еще один элемент нестабильности, который находится в транслируемой области.
Содержание свободного железа в клетке зависит не только от количества в клетке белка ферритина, но и от скорости его поступления в клетку. Скорость поступления железа в клетку возрастает с увеличением на поверхности клетки количества рецепторов для белка трансферина. Последний выполняет функцию переносчика железа. Оказалось, что синтез рецептора трансферина, так же как и ферритина, регулируется железом, но совсем по другому механизму. В присутствии железа в клетке мРНК рецептора трансферина быстро деградирует и синтез этого белка замедляется, в то время как при дефиците железа внутри клетки эта мРНК стабилизируется, обеспечивая высокий уровень синтеза белка. Нестабильность мРНК рецептора трансферина в присутствии железа определяется пятью шпилечными структурами в 3'-НТО (рис. 4, б ). Эти шпилечные структуры (элементы нестабильности мРНК рецептора трансферина) очень похожи на регуляторный элемент в 5'-НТО мРНК ферритина. Элементы нестабильности мРНК рецептора трансферина, так же как регуляторный элемент мРНК ферритина, связывают аконитазу в форме, не содержащей железа. В комплексе с аконитазой мРНК стабилизируется и активно транслируется. В результате на поверхности клеток увеличивается количество рецепторов трансферина и повышается поступление железа в клетки. При увеличении содержания свободного железа в клетках оно связывается с аконитазой. Аконитаза диссоциирует из комплекса с мРНК. Молекула мРНК дестабилизируется и быстро распадается. Это ведет к снижению скорости синтеза рецептора трансферина, уменьшению количества рецепторов на поверхности клетки и замедлению поступления железа в клетку.
Таким образом, один и тот же клеточный белок - аконитаза выполняет в клетке различные функции: в железосодержащей форме катализирует одну из реакций цикла Кребса - обратимое превращение лимонной кислоты в изолимонную, в форме без железа связывается с регуляторным элементом в 5'-НТО мРНК ферритина и репрессирует ее трансляцию, а также с элементами нестабильности в 3'-НТО мРНК рецептора трансферина и защищает эту мРНК от деградации.
СПЕЦИФИЧЕСКИЕ ПОСЛЕДОВАТЕЛЬНОСТИ В мРНК ОТВЕЧАЮТ ЗА ЕЕ ВНУТРИКЛЕТОЧНУЮ ЛОКАЛИЗАЦИЮ
Разные мРНК в эукариотических клетках распределяются в них неодинаково, что особенно легко наблюдать в крупных зародышевых клетках. Наиболее хорошо изучено распределение мРНК в ооцитах плодовой мушки дрозофилы . Так, мРНК, синтезируемые на генах oskar, nanos и bicoid, несколько раз меняют положение в клетке после своего синтеза, что контролируется комплексом генов, пока, наконец, не займут своего окончательного положения: мРНК oskar и nanos на заднем, а мРНК bicoid на переднем конце яйца. Правильное положение этих мРНК задается специфическими последовательностями в их 3'-НТО. Интересно, что сигналы внутриклеточной локализации в 3'-НТО очень протяженные. Вероятно, они формируют сложные вторичные структуры. За окончательную локализацию мРНК oskar и bicoid отвечает белок - продукт гена staufen, узнающий специфическую двухспиральную структуру в 3'-НТО этих мРНК. Интересно, что активная трансляция мРНК oskar и bicoid происходит только в местах их специфической локализации, а их делокализация приводит к подавлению трансляционной активности за счет связывания со специфическими репрессорными белками.