Страница
2
В молекуле феофитина сравнительно легко ввести обратно какой-нибудь металл и восстановить металлоорганическую связь. Для этого к раствору феофитина прибавляют уксуснокислую медь или уксуснокислый цинк и нагревают. Цинк или медь входят в молекулу хлорофилла, и вытяжка становится опять зелёного цвета.
Химическая формула была установлена в 1913 году немецкими биохимиками Р. Вильштеттером и А. Штоллем. Им удалось её установить, последовательно отщепляя от молекулы хлорофилла отдельные её части действием кислот и щелочей, а в дальнейшем и нагреванием под давлением. До этих работ в физиологии растений считалось, что хлорофилл содержит железо, а не магний. Они же окончательно доказали и наличие двух хлорофиллов – а и b.
Эти же работы сделали образование кристаллического хлорофилла. Вильштеттер и Штолль показали, что имеющийся в зелёных листьях фермент хлорофиллаза отщепляет спирт фитол и на его место становится остаток этилового или метилового спирта. Такие соединения получили название хлорофиллидов. Если фитол замещается остатком этилового спирта, то полученное соединение называется этилхлорофиллидом.
Оптические свойства хлорофилла.
Хлорофилл поглощает солнечную энергию и направляет её на химические реакции, которые не могут протекать без энергии, получаемой извне. Раствор хлорофилла в проходящем свете имеет зелёный цвет, но при увеличении толщины слоя или концентрации хлорофилла он приобретает красный цвет.
Хлорофилл поглощает свет не сплошь, а избирательно. При пропускании из семи видимых цветов, которые постепенно переходят друг в друга. При пропускании белого света через призму и раствор хлорофилла на полученном спектре наиболее интенсивное поглощение будет в красных и сине-фиолетовых лучах. Зелёные лучи поглощаются мало, поэтому в тонком слое хлорофилл имеет в проходящем свете зелёный цвет. Однако с увеличением концентрации хлорофилла полосы поглощения расширяются (значительная часть зелёных лучей также поглощается) и без поглощения проходит только часть крайних красных. Спектры поглощения хлорофилла а и b очень близки.
В отражённом свете хлорофилл, кажется вишнёво-красным, так как он излучает поглощённый свет с изменением длины его волны. Это свойство хлорофилла называется флюоресценцией.
Биосинтез хлорофилла
Исходными для синтеза хлорофилла субстратами являются очень простые органические соединения – ацетат и глицин. Процесс синтеза хлорофилла принято подразделять на три этапа.
Первый этап состоит из следующих реакций:
1. Образование ацетилкофермента А, в котором принимает участие ацетат, кофермент А и АТФ. Реакция катализируется ацилкофермент А-синтетазой.
2. Образование сукцинилкофермента А из двух молекул ацетилкофермента А. Считается не менее вероятным другой путь: вовлечение ацетата в цикл Кребса и образование в нём сукцината и затем сукцинилкофермента А. Некоторые исследователи считают исходным субстратом биосинтеза хлорофилла именно сукцинилкофермент А, не рассматривая реакции его образования (как не специфичные, осуществляющиеся в связи и с другими метаболическими цепочками).
3. Образование -амино--кетоадипиновой кислоты из сукцинилкофермента А и глицина, катализируемого так же, как и следующая реакция, ферментом синтетазой -аминолевулиновой кислоты:
4. Из -амино--кетоадипиновой кислоты путём декарбоксилирования образуется -аминолевулиновая кислота:
5. Синтез из двух молекул -аминолевулиновой кислоты пирроленинового кольца и затем изомеризация его в пиррольное кольцо с образованием порфобилиногена. Реакция катализируется ферментом дегидразой -аминолевулиновой кислоты.
Второй этап включает реакции синтеза из четырёх пиррольных колец одной молекулы протопорфирина.
6. Из четырех молекул порфобилиногема под влиянием фермента порфобилиноге-дезаминазы синтезируется цепочка тетрапиррана.
7. Наименее изучен механизм реакции замыкания открытой цепи тетрапиррана в уропорфириноген III.
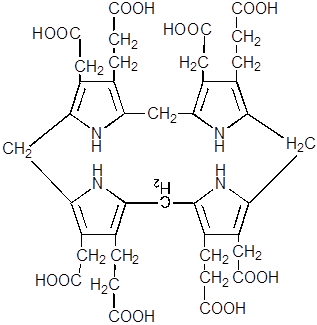 |
8. В результате декарбоксилирования всех четырёх остатков ацетата из уропорфириногена III образуется копропорфириноген III, фермент – уропорфириногендекарбоксилаза.
9. Происходит декарбоксилирование и дегидрирование двух из четырёх пропионатных остатков, что приводит к появлению винильных радикалов в кольцах и образованию протопорфириногена IX, фермент – копропорфириногендекарбоксилаза.
10.
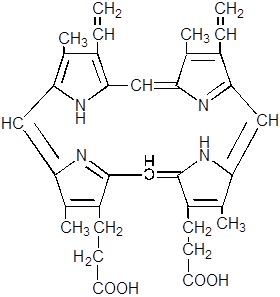 |
Третий этап характеризуется образованием и превращением магнийпорфиринов.
11. Протопорфирин, взаимодействуя с магнием, превращается в магнийпротопорфирин.
12.
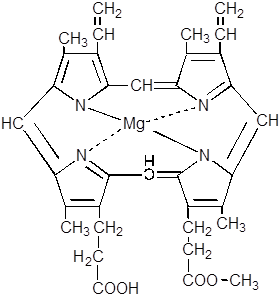 |
13.
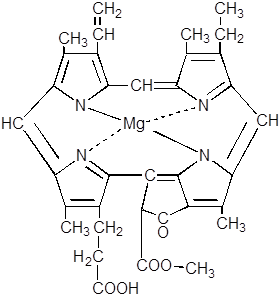 |
14.
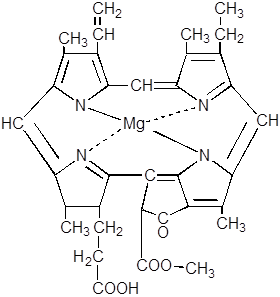 |
Лишь у некоторых низших и голосеменных растений эта реакция может протекать ферментативно в темноте. В хлорофиллид превращается не свободная форма протохлорофиллида, а связанная с белком в единый комплекс – так называемый протохлорофиллидголохром.
15. Последняя реакция – ферментативная этерификация хлорофиллида фитолом, в результате чего образуется хлорофилл а.
Поскольку синтез хлорофилла – многоэтапный, в нём участвуют различные ферменты, составляющие, по-видимому, полиферментный комплекс. Интересно заметить, что образование многих из этих белков-ферментов ускоряется на свету. Содержание хлорофилла в листе колеблется незначительно. Это связано с тем, что идёт непрерывный процесс разрушения старых молекул и образование новых молекул хлорофилла. Причём эти два процесса уравновешивают друг друга. При этом предполагается, что вновь образовавшиеся молекулы хлорофилла не смешиваются со старыми и имеют несколько иные свойства.