Фенологическое картирование в изучении миграций птиц
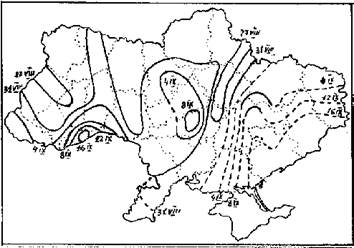
Рис. 3. Карта начала осенней миграции белого аиста на территории Украины (по: Грищенко, Серебряков, 1992).
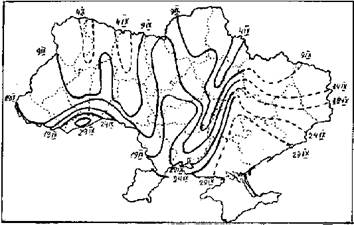
Рис. 4. Карта окончания осенней миграции белого аиста на территории Украины (по: Грищенко, Серебряков, 1992).
Наличие выступов изофен на фенологических картах миграции оказалось всеобщей закономерностью. Они есть на всех картах, построенных нами и В.В. Cеребряковым для Украины (более 20 видов птиц), как для весенней миграции, так и для осенней, как для начала пролета, так и для его окончания. Хорошо видны петли и выступы изофен и на крупномасштабных картах других авторов (Хомченко, 1930, 1972; Beklova et al., 1983 и др.). На картах А.Ф. Миддендорфа, Д.Н. Кайгородова, У. Сливинской, Х. Саутерна, В. Кука изофены идут более или менее плавно. Но в этом случае мы, собственно говоря, уже имеем дело с рассмотрением явления на другом уровне - территория исследуемого региона у названных авторов очень большая: соответственно Российская Империя, ее Европейская часть, Европа, Северная Америка. Понятно, что при построении изофен отдельные выступы сглаживаются. Так, на карте прилета пеночки-веснички (Phylloscopus trochilus) в Европу (Southern, 1938b) вся Украина оказывается между двумя изофенами 1 и 2.04. Подобно этому поверхность листа кажется нам гладкой, но под микроскопом обнаруживаем его сложную структуру. При переходе к более детальным картам появляются выступы изофен. Именно так получилось при обработке фенологического материала за весну 1924 г., когда кроме общих карт были построены и более детальные для отдельных регионов СССР (Святский, 1924б). Для скворца (Sturnus vulgaris) нами была построена фенокарта прилета на юге Западной Украины (Грищенко, 1992). Она показала дробление единого миграционного потока, проходящего на северо-восток через Карпаты, на более мелкие, т. е. можно говорить о миграционных потоках второго порядка, существующих внутри более крупных.
Неравномерность наступления сроков миграции - объективно существующее явление. Выпячивания изофен, охватывающие территории с более ранними сроками миграции, мы предлагаем называть фенологическими руслами пролета. Термин "миграционный поток" неплохо отражает суть явления, но он соответствует действительности лишь для карт начала миграции, выпячивания изофен же есть и на картах последнего наблюдения. Термин же "пролетный путь" имеет слишком одиозную историю, разные авторы дают ему различную трактовку, наконец соответствие фенологических русел пролета пролетным путям в современном их понимании, собственно говоря, еще предстоит доказать, то есть убедиться в более высокой интенсивности миграции на их территории. А это возможно лишь при привлечении большого количества наблюдателей, проведении радарных наблюдений и т. п. Но хотя непосредственные доказательства такого соответствия представить сложно, есть немало косвенных. Размещение фенологических русел пролета на Украине совпадает с ходом пролетных путей, установленным другими методами (по: Браунер, 1923; Штейнбахер, 1956; Флинт, Панчешникова, 1982, 1985 и др.). Они совпадают для различных экологически близких видов, а известно, что общие пути пролета и места зимовки свойственны не только популяции одного вида, а и популяциям птиц со сходными экологическими требованиями, гнездящимся в одном географическом регионе (Михеев, 1978). Наконец, вполне логично ожидать появление первых мигрантов на наиболее "проторенных дорожках" - пролетных путях.
Структурированность миграционного ареала характерна не только для начала пролета, она остается более или менее постоянной. Нашими исследованиями установлено почти полное соответствие фенокарт начала и окончания осенней миграции для всех четырех видов, по которым мы имеем такие данные: белого аиста, серого журавля, гусей, кряквы. Лучше всего это проявляется у белого аиста - фенологические русла в начале и в конце пролета совпадают практически полностью (рис. 3, 4). То есть перелет заканчивается раньше там же, где и начинается. Для всех четырех видов существует достоверная положительная корреляция между средними многолетними датами начала и окончания миграции (Грищенко, 1990). У других птиц на фенокартах последнего наблюдения фенологические русла примерно совпадают с таковыми на картах начала миграции иных видов. Очевидно, эта закономерность также есть всеобщей. Размещение фенологических русел на протяжении всей миграции сохраняется, они могут лишь несколько смещаться или отклоняться. То, что птицы раньше заканчивают миграцию на пролетных путях, можно объяснить большей ее интенсивностью там. Это хорошо видно и на картах Расстояние между изофенами на фенологических руслах, как правило, больше, чем за их пределами, и значительно больше, чем в областях запаздывания (см., например, карту прилета снегиря (Pyrrhula pyrrhula) (Грищенко, Серебряков, 1991), то есть фронт миграции здесь продвигается быстрее.
Сейчас мы уже знаем, что птицы осенью не "удирают" от холода, бескормицы и т. п. Во время перелета у них развивается особое миграционное состояние. Птицы с высокоразвитым миграционным состоянием будуть продолжать перелет, какими бы благоприятными ни были условия в местах остановки (Дольник, 1975). Одним из важнейших элементов миграционного состояния является наличие жировых резервов. Есть некоторые свидетельства, что в местах более раннего появления у птиц эти резервы выше. Так, в окрестностях Горького и Перми у 1964 г. проводился отстрел чечеток (Acanthis cannabina) с целью анализа их жировых запасов. В Перми эти птицы появляются на две недели раньше и здесь же, как оказалось, они имеют большее количество жира (Постников, 1970). По данным Ю.Я. Руте (1976), скорость миграции синиц тем больше, чем выше ее интенсивность. Интенсивность миграции наибольшая на пролетных путях, этим и объясняется более раннее появление там птиц. Благодаря постоянству размещения фенологических русел пролета для выявления пролетных путей можно использовать карты не только начала миграции, а и последнего наблюдения.
Фенологические карты миграции несут довольно большую информацию. Простое сравнение сроков прилета или отлета для разных частей изучаемого региона становится более наглядным, при этом можно судить о ходе миграционного продвижения птиц. Наконец, фенокарты отображают структуру миграционного ареала. Что ценно, они дают возможность проводить анализ закономерностей на различных уровнях, в зависимости от размеров изучаемой территории и степени сглаживания линий.
Это можно хорошо проиллюстрировать на примере проблемы, которая вызывала многочисленные споры - соответствие сроков прилета птиц ходу весны, прежде всего температуре в местах наблюдения. Еще Д.Н. Кайгородов (1908) нанес на карту не только изохроны прилета кукушки, а и изотермы. Оказалось, что эти линии идут примерно параллельно. Параллельны изотермам изофены ряда видов и на картах Х. Саутерна и В. Кука. Можно сделать вывод о тесной связи между прилетом птиц и ходом весны. Но уже К. Бречер (1916b) и В. Экардт (1929) указывают на основании анализа наблюдений в отдельных пунктах, что такой связи нет, прилет от температуры не зависит. Позже этот же тезис развивают Н.А. Гладков (1937) и В.Ф. Гаврин (1957). Кто же прав? По нашему мнению, в данном случае мы имеем дело с закономерностями различных уровней. Фронт миграции в целом продвигается примерно в соответствии с изменением температуры, но уже на уровне отдельных регионов эта закономерность не оправдывается, прежде всего из-за структурированности миграционного ареала - наличия фенологических русел пролета и областей запаздывания.