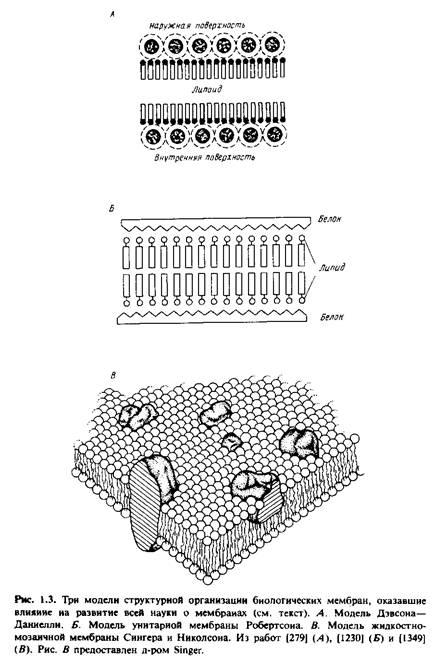
Структура и состав биологических мембран
Ядерная мембрана. Ядерная оболочка клетки, находящейся в интерфазе, на электронных микрофотографиях имеет вид двух элементарных мембран с узким просветом между ними, называемым перинуклеарным пространством. Эта мембрана происходит из эндоплазматического ретикулума и, по-видимому, неразрывно связана с ним. Наиболее характерными морфологическими признаками являются порообразные структуры. Они имеют диаметр около 600 А и состоят из морфологически четко выявляемых компонентов, образующих октагональную решетку. В том месте, где расположены эти структуры, внутренняя и наружная ядерные мембраны выглядят слившимися. Полагают, что поры позволяют комплексам мРНК-белок переходить из ядра в цитоплазму, а регуляторным белкам перемещаться в обратном направлении, из цитоплазмы в ядро. Биохимические данные о ядерной оболочке весьма немногочисленны.
Эндоплазматический ретикулум. Это сложная сеть цистернообразных или трубчатых структур, которая занимает значительную часть внутреннего объема обычной животной клетки. Основная роль ЭР состоит в том, что он служит местом биосинтеза белков, которые затем секретируются, включаются в лизосомы или в плазматическую мембрану. Потенциально опасные для клетки гидролитические ферменты, которые должны секретироваться или накапливаться в лизосомах, подвергаются в ЭР процессингу до зрелой формы. С ЭР часто бывают связаны рибосомы, в результате чего на электронных микрофотографиях он выглядит шероховатым. Сложные процессы, в ходе которых осуществляется синтез белков, их превращение в зрелую форму и направленная доставка к месту назначения, описаны в гл.10.
Области ЭР, не содержащие рибосом, называются гладким ЭР. Здесь осуществляется биосинтез стеролов, протекают реакции детоксикации и происходит десатурация жирных кислот. Все эти процессы входят в сложную, согласованную систему транспорта электронов, осуществляемого при участии цитохромов bs и Р450.
Аппарат Гольджи. Эта органелла состоит из сети уплощенных мешков, собранных в стопки. Основная его функция заключается в посттрансляционной модификации гликопротеинов, синтезированных в эндоплазматическом ретикулуме и предназначенных для секреции, включения в плазматическую мембрану или доставки в лизосомы. Эти органеллы содержат гликозидазы и гликозилтрансферазы, которые вступают в действие последовательно, по мере того как белок, подвергаемый процессингу, перемещается от начала аппарата Гольджи до его конца. Фактически аппарат Гольджи состоит из совокупности отдельных мембран, образующих цистерны. Эти мембраны, которые можно выделить, характеризуются определенным набором ферментов. Механизмы транспорта мембран и секретируемых белков через аппарат Гольджи рассмотрены в гл.10.
Лизосомы. Эти органеллы ответственны за деградацию макромолекул и содержат ряд гидролитических ферментов, таких, как протеазы и липазы. Вещества, захваченные клеткой путем эндо- или фагоцитоза, которые необходимо расщепить, доставляются в лизосомы с помощью везикул. В лизосомах происходит также расщепление клеточных компонентов в ходе их обычного круговорота. Как осуществляются синтез лизосомных ферментов, их маркировка для доставки в лизосомы и последующий транспорт - изучено достаточно хорошо. Эти процессы рассматриваются в гл.10.
Пероксисомы. Эти органеллы содержат окислительные ферменты, участвующие в деградации малых молекул, таких, как аминокислоты, ксантин и, в особенности, жирные кислоты. Их название связано с присутствием в них каталазы, которая разлагает перекиси, образующиеся как побочные продукты при реакциях окисления.
Митохондрии. В этих органеллах осуществляется окислительное фосфорилирование, в результате чего в ходе окисления субстратов, таких, как NADH или сукцинат, образуется АТР. Митохондрии образованы двумя мембранами, разделенными некоторым промежутком. Внутренняя область митохондрий называется матриксом. Внутренняя мембрана образует складки в виде перегородок, называемых кристами, и содержит ферменты, участвующие в транспорте электронов и синтезе АТР. В гл.6 обсуждаются роль диффузии в плоскости мембраны компонентов цепи электронного транспорта и ее функциональное значение. Вопросы, связанные с синтезом белков митохондрий, который происходит в цитоплазме, и с их доставкой в один из митохондриальных компартментов или в одну из мембран, рассматриваются в гл.10.
Хлоропласты. Это органеллы, содержащие фотосинтетический аппарат. Они имеют наружную оболочку, образуемую двумя мембранами, и внутреннюю область - строму. В строме находятся тилакоидные мембраны, где локализованы компоненты системы фотосинтеза. На отдельных участках тилакоидные мембраны плотно упакованы в стопки, а на других обращены непосредственно к строме. Состав плотноупакованных и обращенных в строму доменов тилакоидной мембраны различен, что указывает на латеральную гетерогенность этой мембраны. Энзимология фотосинтезирующей цепи электронного транспорта обсуждается в разд.6.6.
Следует подчеркнуть, что при изучении каждой из мембран, упомянутых выше, а также и других специализированных мембран из клеток животных, растений или бактерий возникает целый комплекс важных и интересных вопросов, требующих своего решения, и открываются широкие возможности для биохимических исследований. Другие мембранные системы, представляющие интерес в этом отношении, будут описаны в последующих главах книги.
2. Исторический очерк
Тот факт, что плазматическая мембрана, окружающая клетки, представляет собой вполне определенную структуру, был осознан в середине XIX столетия.
На исходе этого столетия Овертон обратил внимание на корреляцию между скоростью, с которой небольшие молекулы проникают в растительные клетки, и их коэффициентом распределения между маслом и водой; это привело его к мысли о липидной природе мембран. В 1925 г. Гортер и Грендел предположили, что липиды в мембране эритроцитов образуют биомолекулярный слой. Эта идея возникла на основе результатов элегантного и простого эксперимента.
Липиды эритроцитов экстрагировали ацетоном и затем в кювете Лэнгмюра получали из них тонкую пленку на поверхности воды.
С помощью поплавка сжимали слой липидных молекул на границе раздела вода-воздух до тех пор, пока этот слой не начинал оказывать сопротивление дальнейшему сжатию; это явление было объяснено образованием плотноупакованной мономолекулярной липидной пленки. Измерение площади, занимаемой липидами, и сравнение ее с площадью поверхности эритроцитов, из которых эти липиды были экстрагированы, дали соотношение 2:
1. Отсюда был сделан вывод, что мембрана эритроцитов состоит из липидных молекул, расположенных в два слоя.
По-видимому, этот вывод Гортера и Грендела оказался правильным только благодаря взаимной компенсации ошибок, однако в историческом плане эта работа имела большое значение, поскольку с тех пор концепция липидного бислоя как структурной основы биологических мембран стала доминирующей и на самом деле оказалась верной.