Строение и состав живой клетки

Кроме фосфолипидов, построенных на основе глицерина, в клетках эу- и прокариот встречаются фосфолипиды, которые являются производными диодов: этиленгпиколя, 1,2- и 1,3-пропандиолов, 1,3-, 1,4- и 2,3-бутандиолов, а также 1,5-пентандиола. Среди диольных липидов встречаются моно- и диацильные производные — это сложные эфиры различных жирных кислот; простые эфиры, смешанные алкильные (или алкенильные) и ацильные производные, диольные аналоги фосфатидилхолина, фосфатидилэтаноламина и др. Обычно содержание диольных липидов составляет только 0,5—1,5% содержания глицериновых липидов. Но некоторые морские моллюски и иглокожие содержат в тканях примерно равные количества производных глицерина и этиленгликоля. Содержание диольных липидов уменьшается в течение зимы, поэтому они предположительно могут использоваться как запасные вещества. Замечено также их увеличение в процессе регенерации печени у крыс.
Другим классом «безглицериновых» липидов являются сфинголипиды, основу которых составляет алифатический аминоспирт сфингозин (или дигидросфингозин) (рис. 7). Построенные на его основе церамиды широко распространены в тканях растений и животных, но количество их незначительно. Они обнаружены также в пилях Escherichia coli. Сфингомиелины входят в состав нервной ткани, липидов крови и некоторых других компонентов клеток животных.

В большинстве биологических мембран содержатся также гликолипиды. В клетках животных они, как правило, являются производными сфингозина, у которого к первичному гидроксилу присоединен остаток сахара или олигосахарида. Если сахарами являются глюкоза или галактоза, такие липиды называются цереброзидами (особенно много их в тканях мозга). Если углеводной частью является олигосахарид, речь идет о ганглиозидах (выделены из ганглий, мозга и ряда других тканей). Ганглиозиды найдены также в пилях некоторых бактерий (Neisseria) и определяют прилипание этих патогенов к поверхности клеток животного организма. У прокариот (в основном у грамположительных бактерий и цианобактерий) гликолипиды также содержат глюкозу, галактозу и маннозу. Их количество в мембранах невелико, но при лимитировании фосфором может возрастать, при этом гликолипиды, по-видимому, замещают фосфолипиды. Наконец, в мембранах большинства эукариот (а также у Mycoplasma) содержатся стероиды, в основном холестерин (у животных), эргостерин (у дрожжей) и стигмастерин (у растений). Микоплазмы неспособны синтезировать стерины, но требуют присутствия их в среде для стабилизации клеточной мембраны.
3. Природа и состав жирных кислот в мембранных липидах
Природа жирных кислот в липидах мембран зависит как от вида организма, так и от условий его существования. Наиболее часто встречающиеся жирные кислоты липидов животных, растительных и прокариотных клеток.
С увеличением числа двойных связей значительно снижается температура плавления жирных кислот (а также содержащих эти кислоты липидов) и повышается их растворимость в неполярных растворителях. Поскольку функциональная активность мембранных белков регулируется фазовым состоянием липидов мембраны (как правило, в жидком состоянии их активность выше), при снижении температуры в мембране должно повышаться содержание ненасыщенных кислот. Благодаря постоянству внутренних условий (гомеостазу) животного макроорганизма влияние температуры на жирнокислотный состав липидов обычно проявляется слабо, но, например, в липидах нижних конечностей пингвинов повышено содержание ненасыщенных жирных кислот.
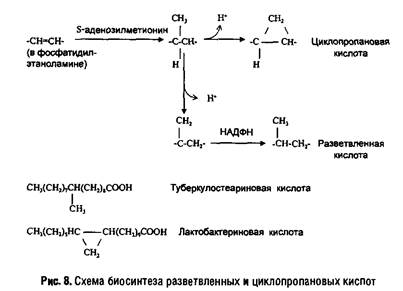
В мембранах прокариот разнообразие жирных кислот довольно велико. Особенностью бактерий является наличие разветвленных и циклопропановых кислот. У некоторых грамположительных бактерий (например, Micrococcus luteus) 90% жирных кислот липидов составляют разветвленные кислоты, а ненасыщенные кислоты практически отсутствуют. Грамотрицательные бактерии содержат смесь насыщенных и ненасыщенных кислот с преобладанием С-16 и С-18 кислот, а также циклопропановые кислоты (например, лактобацилловую кислоту). Пути их биосинтеза показаны на рис. 8.
Изменение температуры существенно влияет на соотношение насыщенных и ненасыщенных жирных кислот в мембранах прокариот, при этом снижение температуры культивирования приводит к увеличению доли ненасыщенных кислот. Предельными случаями являются психрофилы, растущие при температурах, близких к 0"С, у которых присутствуют практически только ненасыщенные жирные кислоты, а также термофилы (привычная температурная среда выше 60°С) — у них все жирные кислоты липидов насыщенные.
Тем не менее состав жирных кислот фосфолипидов у бактерий при культивировании в стандартных условиях достаточно постоянен, и его можно использовать как таксономический признак — в интересах классификации или при анализе состава микробных сообществ без изолирования в чистых культурах отдельных представителей.
Для изучения структуры, состава и функции органелл необходимо, как правило, изолировать их из клетки в чистом виде. Обычно это достигается методом дифференциального центрифугирования после разрушения клеток в механических или ультразвуковых гомогенизаторах.
Кратко охарактеризуем основные процессы и органеллы (в алфавитном порядке).
Аэросомы — однослойные везикулы, мембрана которых построена только из белка. Способствуют повышению плавучести клеток, так как в них содержится газовая фаза, совпадающая по составу с газовой фазой окружающей среды.
Вакуоли — мембранные образования, служащие для поддержания тургорного давления, запасания различных веществ, а также выполняющие лизосомные функции.
Внехромосомная ДНК. В митохондриях и хлоропластах содержится ДНК, образующая нуклеоид бактериального типа. Заключенная в ней генетическая информация не дублируется в ядерной ДНК и способна к автономному выражению в белках посредством собственных систем транскрипции и трансляции (включающих рибосомы 70 S бактериального типа).
У прокариот внехромосомная ДНК организована в виде плазмид, которые могут существовать и реплицироваться автономно или в интегрированном в хромосому состоянии (например, в виде профага).
Гидрогеносомы — окружены однослойной мембраной и содержат комплекс пируватдегидрогеназ (у трихомонад).
Гистоны — представляют собой положительно заряженные (основные) белки, входящие в состав хромосом в комплексе с ДНК (обнаружены также у архебактерий).
Гликосомы — окружены однослойной мембраной и содержат ферменты гликолиза (у некоторых протозойных микроорганизмов, в частности у возбудителей сонной болезни).
Глиоксисомы — разновидность пероксисом — место локализации ферментов глиоксалатного шунта, участвующих в превращении запасных жиров в углеводы. Поэтому они тесно ассоциированы со сферосомами, жирозапасающими органеллами растительных клеток.
Жгутики и реснички представляют собой аппарат, определяющий подвижность клеток или их способность создавать поток окружающей среды к органам поглощения пищи. Жгутики эукариот и прокариот сильно различаются по строению и составу.