Страница
2
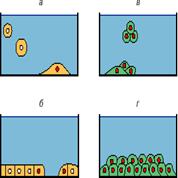
Рис. 3. Регуляция размножения клеток в культуре. Жидкая среда синяя, стенки и дно сосуда черные. У неразмножающихся клеток ядра не окрашены, у клеток в митотическом цикле красные: а, б - нормальные клетки: а - зависимость размножения клетки от подложки: вступила в цикл лишь клетка, прикрепившаяся к поверхности дна сосуда (подложке), но не клетки, взвешенные в жидкой среде, б - зависимость размножения от плотности населения культуры. Среди клеток, прикрепленных к подложке, размножаются лишь те, которые граничат со свободной поверхностью этой подложки, но не клетки, окруженные со всех сторон соседями; в, г - опухолевые клетки размножаются без подложки в жидкой среде (в) и на подложке независимо от числа соседей
В отличие от контактов с подложкой контакты с другими клетками, по-видимому, приводят к торможению размножения (рис. 3). Действительно, когда в результате размножения клеток культура становится густой и на ее дне не остается незанятых клетками мест, то эти клетки перестают делиться, то есть выходят из митотического цикла в так называемую фазу покоя Go [5]. Если сделать рану (удалить бритвой со дна сосуда небольшой участок густой культуры), то контактное торможение на краю раны исчезает и клетки начинают мигрировать в рану на свободную подложку. Вскоре такие мигрировавшие клетки начинают в ране активно размножаться и делятся до тех пор, пока не зарастет вся рана, то есть свободная подложка. Эти опыты свидетельствуют о том, что размножение зависит от местной "густоты" населения культуры. Хотя они не доказывают строго, что именно прямые контакты клетка-клетка тормозят размножение, такая возможность представляется очень вероятной.
Контроль размножения с соседними клетками и подложкой в культуре аналогичен процессам, происходящим в организме, Приведем только один пример. Представьте себе царапину, нарушившую целостность эпителия кожи. Клетки эпителия по краю царапины начинают мигрировать на поверхность свободного матрикса и размножаются до тех пор, пока эта поверхность будет полностью закрыта.
Во всех случаях регулируется местное соотношение клеток и территорий, на которых они находятся. Нормальная клетка размножается только тогда, когда у нее есть свободная "площадь для жилья". Благодаря этим регуляциям поддерживается и восстанавливается после повреждений правильная тканевая структура (заживление ран!).
НАРУШЕНИЯ МОРФОГЕНЕТИЧЕСКИХ РЕАКЦИЙ ПРИ ОПУХОЛЕВЫХ ТРАНСФОРМАЦИЯХ
Опыты с культурами показали, что при опухолевых трансформациях эпителиоцитов и фибробластов резко нарушаются все морфогенетические реакции на контакты с другими клетками и подложкой. Такие трансформированные клетки делают меньше матрикса, хуже прикрепляются к матриксу, подложке и другим клеткам. Нарушаются также контактные регуляции размножения. Например, клетки, взвешенные в вязкой среде, продолжают размножаться без прикрепления к подложке. Эти клетки размножаются и в густой культуре независимо от числа соседей. В результате тканевые системы эпителия и соединительной ткани в культуре и организме становятся хуже организованными, а в предельном случае такие системы вообще не образуются, но распадаются на отдельные клетки и группы клеток. В организме такие отделившиеся от ткани клетки начинают двигаться на соседние территории и размножаться там. Например, трансформированные клетки эпителия, отделившись от пласта, начинают мигрировать через дефектную базальную мембрану в окружающую соединительную ткань, а затем проникать в просвет сосудов, откуда они током лимфы или крови могут переноситься в другие органы. Такие крайние изменения способности к морфогенетическим реакциям и построению тканей обусловливают наиболее опасные свойства опухолевых клеток - способность к врастанию в нормальные ткани (инвазия) и к образованию колоний в других органах (метастазирование). Отметим, что способность к инвазии и метастазированию свойственна лишь клеткам опухолей, наиболее резко измененным генетически. Врачи называют такие опухоли злокачественными (рис. 4). Клетки с менее измененными морфогенетическими реакциями образуют доброкачественные опухоли, где имеются различные нарушения организации тканевых структур, но нет инвазии и метастазирования. Анализ генома показал, что клетки злокачественных опухолей человека имеют целую серию мутаций разных онкогенов и антионкогенов, тогда как клетки доброкачественных опухолей обычно имеют меньшее число таких мутаций. Таким образом, степень нарушения морфогенетических реакций и соответственно течение опухоли определяется числом и степенью изменений генома опухолевых клеток, одной мутации для развития рака обычно недостаточно.
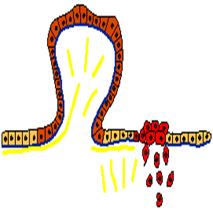
Рис. 4. Два опухолевых клона среди клеток нормального эпителия. Нормальные клетки светло-коричневые. Клетки клона, образующего доброкачественную опухоль - полип (темно-коричневый цвет), дают избыточный рост и образуют выпячивание над поверхностью пласта, но не проникают в соединительную ткань под эпителием. Напротив, клетки злокачественного (ракового) клона (красный цвет) проникают под базальную мембрану в соединительную ткань
МЕХАНИЗМ РЕГУЛЯЦИИ КОНТАКТНЫХ РЕАКЦИЙ
В первой части статьи [5] мы разобрали основы молекулярных механизмов действия специальных сигнальных молекул из жидкой среды, регулирующих размножение клеток. Напомним, что такие молекулы связываются со специальными рецепторами в наружной мембране клетки и активируют эти рецепторы, а те, в свою очередь, активируют цепь промежуточных белков в цитоплазме, что в конце концов приводит к активации синтеза ядерных белков, необходимых для удвоения генома (ДНК) и подготовки деления. Онкогены опухолевых клеток - мутировавшие гены, кодирующие белки разных этапов цепей проведения сигналов. При опухолевых трансформациях онкогены, вызывающие нарушения реакций на гуморальные факторы, индуцируют, как мы видели, и нарушения морфогенетических реакций. Отсюда следует, что многие этапы обеих групп реакций на сигналы контролируют одни и те же белки, в частности специальные ферменты, присоединяющие фосфат к белку (киназы, см. [5]). Конечными этапами цепей активации морфогенетических реакций становятся, вероятно, какие-то белки цитоскелета, например, белки, связывающиеся с актиновыми филаментами [3] и вызывающие изменения сборки и сокращения псевдоподий и образования контактных адгезий. Действительно, у некоторых таких белков присоединение фосфата резко повышено в трансформированных клетках, однако пока еще неясно, каково значение изменений каждого конкретного белка в нарушениях морфогенеза. Другой нерешенный вопрос - какие рецепторы определяют контактную регуляцию размножения нормальных клеток. Вероятное предположение состоит в том, что активация и торможение размножения зависят от тех же белков, которые прикрепляют клетку к подложке и другим клеткам, то есть от белков-рецепторов мембраны адгезионных структур, соединяющихся с белками подложки или соседних клеток. Оказалось, что адгезионные структуры с подложкой (так называемые фокальные контакты, рис. 5) содержат не только белки, необходимые для механического соединения белков матрикса через мембрану с цитоскелетом, но и несколько десятков видов других белков, функции которых неясны. Среди этих "непонятных" белков имеются различные протоонкобелки, участвующие в проведении сигналов, в том числе ферменты-киназы. Зачем они здесь? Естественно предположить, что фокальные адгезии не только механические "склейки", но и "органы осязания" клетки, передающие сигнал от соприкосновения с другой поверхностью с наружной мембраны внутрь клетки и вызывающие активацию размножения. Возможно, что рецепторы адгезий клетка-клетка, наоборот, индуцируют другие сигналы, тормозящие размножение. Все эти гипотезы сейчас активно проверяются в разных лабораториях мира. Взаимодействие системы, проводящей сигналы от мембраны, с цитоскелетом определяет ту разумную организацию движений и изменений формы клеток, тот "разум цитоскелета", о котором говорилось в предыдущей статье [4].