Сезонная динамика фитопланктона Средней Оби в 2007-2008 гг.
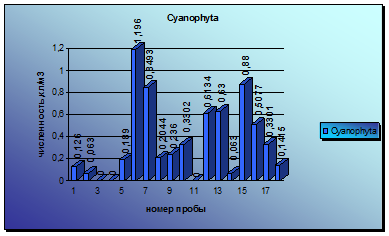
Анализируя колебания численности зеленых микроводорослей, можно отметить 3 небольших и один крупный всплеск вегетации водорослей. Первый подъем наблюдается уже в середине мая, затем уровень вегетации немного снижается до следующего пика численности в середине июля. Первый подъем был вызван водорослями родов Closterium и Pleurotaenium, второй - развитием водорослей родов Closterium и Palmella. Затем наблюдается медленный и постепенный спад вегетации, который сходит почти до нуля. Но в середине августа наблюдается резкий скачок численности, который постепенно к концу сентября переходит в еще один всплеск численности, самый крупный за весь период наблюдений вегетации всех отделов водорослей. Он был вызван в основном развитием водорослей р. Ankistrodesmus и Closterium (диаграмма 4).
Диаграмма 4 - Сезонная динамика Chlorophyta в 2008 г.
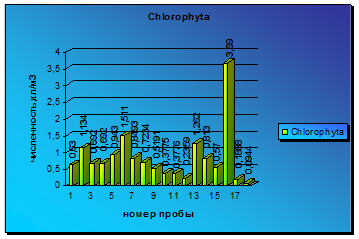
Колебания численности диатомовых проявляются хорошо и периодически снижаются до нуля. Из 18 проанализированных проб в 7 не было обнаружено водорослей данного отдела. Достаточно высокий уровень вегетации диатомовых был зарегистрирован в июньских пробах. В этот период наблюдалась вегетация видов Asterionella formosa Hass, Fragillaria construens (Ehr) Grum var. Subsalina Hust, Pinnularia bigibba Ehr. Var. mesogondyla f. interrupta, Synedra tabulata (Ag) Kütz tabulate и Melosira islandica O. Müll subsp. islandica Hust. Далее, с июня по август, встречались лишь отдельные представители отдела диатомовых, не вносящих большого вклада в общий уровень вегетации водорослей. Только в начале сентября вновь повысился уровень вегетации диатомовых. Он продержался на невысоком уровне до конца сентября и вылился в самую крупную вспышку численности диатомовых, вызванную развитием представителей родов Amphora, Opephora, Pinnularia и Navicula (диаграмма 5).
Таким образом, вегетация диатомовых за весь период наблюдений не достигала высоких уровней, а периодами условия были неблагоприятными для развития представителей отдела Bacillariophyta, что вызывало полное их отсутствие в исследуемых пробах.
Часто лимитирующим фактором для диатомовых является содержание в воде Si. По мнению большинства исследователей, для того, чтобы наступило "цветение" водоема за счет бурного развития диатомовых, необходимо содержание кремния в воде не менее 4мг/л. Однако в природе часто и при меньших концентрациях Si диатомовые водоросли продолжают развиваться. По отношению к кислотности среды диатомовые более приурочены к кислым водам (О.Г. Воропаева, 1988г). Но, по-видимому, температурный режим, химический состав воды и другие фактор помешали нормальному развитию водорослей.
Диаграмма 5 - Сезонная динамика Bacillariophyta в 2008 г.
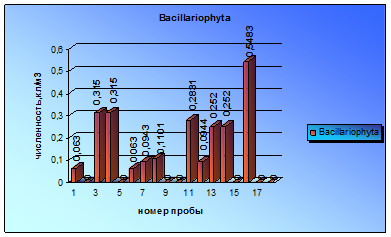
Представители отдела Euglenophyta встречены в пробах еще реже диатомовых. Только в 5 пробах были зарегистрированы небольшие вспышки численности евгленовых. Впервые микроводоросли данного отдела были обнаружены в пробе за 3 июля и периодически встречались до начала сентября (диаграмма 6).
На протяжении этого периода можно отметить незначительные увеличения численности, которые достигли наибольшей величины в пробах за 1 сентября. Максимальный всплеск численности водорослей был вызван интенсивной вегетацией видов Trachelomonas lacustris Drez. Emend Balech и Trachelomonas lacustris var. Sabutata (Skv) Popova.
Видовой состав евгленовых на протяжении всего времени их вегетации постоянно менялся. И невозможно найти одни и те же виды, встречающиеся в разные периоды времени.
Диаграмма 6 - Сезонная динамика Euglenophyta в 2008 г.
6. Определение сапробности исследуемого участка реки Обь в 2008 году
В ходе исследований в 2008 году нами было выявлено 29 видов-индикаторов сапробности. Из них 16 видов зеленых, 7 видов диатомовых, 5 - эвгленовые и 2 вида сине-зеленые. Чаще всего встречались виды, характерные для β-мезосапробной (13 видов) и олигосапробной (9 видов) зон. Обнаружено также 4 вида, характерных для 0-β-сапрбной зоны, 1 вид - для β-α-мезосапробной зоны и 3 вида α-мезосапробной зоны (таблица 10).
Таблица 10 - Виды - индикаторы сапробности.
|
отдел |
вид |
сапробность | |
|
Chlorophyta |
Ulotrix zonata Kűtz |
0 | |
|
Bacillariophyta |
Amphora ovalis Kütz var. pediculus Kütz |
0 | |
|
Bacillariophyta |
Asterionella formosa Hass |
0 | |
|
Bacillariophyta |
Navicula tuscula (Ehr) Grun |
0 | |
|
Bacillariophyta |
Fragillaria construens (Ehr) Grum var. Subsalina Hust |
0 | |
|
Bacillariophyta |
Synedra tabulata (Ag) Kütz tabulata |
0 | |
|
Bacillariophyta |
Nitzschia vermicularis (Kűtz) Grun |
0 | |
|
Cyanophyta |
Gloeocapsa minuta (Kűtz) Hollerb ampl |
0 | |
|
Bacillariophyta |
Nitzschia gracilis Hant |
0 | |
|
Cyanophyta |
Anabaena spiroides Kleb |
0-β | |
|
Chlorophyta |
Scenedesmus acuminatus (Legerh) Chod |
0-β | |
|
Euglenophyta |
Euglena granulata (Klebs) Schmitz |
0-β | |
|
Euglenophyta |
Lepocinclis fusiformes (Carter) Lemm |
0-β | |
|
Chlorophyta |
Crucigenia tetrapedia (Kirhn) W. et. G West |
β | |
|
Chlorophyta |
Gonium sociale Warm |
β | |
|
Cyanophyta |
Microcystis pulverea (Wood) Forti emend f. parasitica (Kűtz) Elenk |
β | |
|
Chlorophyta |
Closterium moniliferum (Bory) Ehr. |
β | |
|
Chlorophyta |
Pandorina morum (Müll) Bory |
β | |
|
Chlorophyta |
Pediastrum simplex Meyen |
β | |
|
Chlorophyta |
Pediastrum dulex Meyen |
β | |
|
Chlorophyta |
Pediastrum tetras var. tetras |
β | |
|
Chlorophyta |
Tetraëdron caudatum (Сorda) Hansg |
β | |
|
Chlorophyta |
Tetraëdron minutum (A. Br) Hansg |
β | |
|
Chlorophyta |
Actinastrum hantschii Lagerh |
β | |
|
Chlorophyta |
Anristrodesmus acicularis Korschik |
β | |
|
Chlorophyta |
Closterium parvulum Näg |
β | |
|
Cyanophyta |
Merismopedia tenuissima Lemm |
β-α | |
|
Cyanophyta |
Oscillatoria splendida Grev |
α | |
|
Chlorophyta |
Closterium acerosum (Schr) Ehr. |
α | |
|
Chlorophyta |
Cosmarium botrytis Menegh |
α | |