Страница
2
Управление ростом аксона
Аксоны нервных клеток могут достигать 1 метра и более в длину, образуя синапсы в определенном месте на строго определенной клетке в области, где имеется большое количество других потенциальных клеток-мишеней. Две основные теории, касающиеся того, каким образом устанавливается такая специфичность в синаптических связях во время развития, были предложены в первой четверти двадцатого века. Согласно одной, нейроны и их мишени заранее запрограммированы на образование определенных синаптических связей. Другая теория считает, что первоначально связи устанавливаются более или менее случайно, а затем часть из них реорганизовывается благодаря влияниям на нейроны со стороны клеток-мишеней, в результате чего происходит устранение ошибочных синапсов и гибель неправильно соединенных клеток. Экспериментальные доказательства, полученные к настоящему времени, говорят в пользу того, что рост аксонов и образование синаптических связей является селективным процессом; аксон направляется строго к своей цели благодаря определенным сигналам в своем окружении.
Навигация аксона, зависящая и не зависящая от клетки-мишени
Какие внеклеточные сигналы управляют конусом роста? Рамон-и-Кахаль сначала предложил хемоаттрактантную модель управления аксоном, согласно которой конусы роста аксонов следуют по градиенту концентрации некоторых молекул, вырабатываемых клетками-мишенями. Такой механизм возможен для роста аксона, когда расстояние между телом нейрона и его мишенью очень короткое. Например, Лумсден и Дэвис изучали рост аксонов из ганглия тройничного нерва, расположенного в голове мыши, в соседние эпителиальные ткани, расстояние до которых составляло менее 1 мм (эти аксоны в конечном счете образовывали сенсорную иннервацию вибрисс, Если развивающийся ганглий тройничного нерва помещался в культуру вблизи эксплантантов из нескольких периферических тканей, нервные отростки росли из ганглия в направлении собственной клетки-мишени, игнорируя другие ткани. Эксплантаты эпителия ткани-мишени оказывали подобное действие на рост аксонов только в том случае, если они брались у эмбриона в период становления иннервации.
В противоположность этому, способность аксонов спинальных мотонейронов расти в направлении конечностей не зависит от наличия мышечной клетки-мишени. Это было показано путем удаления в раннем периоде сомита, из которого развивается мускулатура конечностей. Аксоны мотонейронов направляются нормально из спинного мозга, врастают в конечность и образуют определенный паттерн мышечных нервов, даже в отсутствие мышцы. Таким образом, факторы, которые управляют ростом аксонов мотонейронов к определенной мишени на конечности, не выделяются мышцами, с которыми аксоны в конечном итоге образуют связи.
|
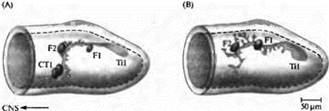
Рис. 4. Конусы роста периферических нейронов используют клетки-ориентиры для навигации в конечностях кузнечика. (А) В нормальном эмбрионе аксоны Ti1 нейрона встречают на своем пути в центральную нервную систему серию клеток-ориентиров: клетки Fl, F2 и две клетки СТ1. (В) Если СТ1 клетку убить на ранней стадии развития, нейрон Ti1 образует несколько аксональных веточек из точки, где находится F2, и конусы роста направляются в неправильном направлении. |
|
Навигация по клеткам-ориентирам (guidepost cells)
Когда дистанция от нейрона до его цели составляет больше чем несколько сотен микрон, его путь обозначен специальными промежуточными целями. Например, конус роста, идущий от сенсорной клетки конечностей у развивающегося кузнечика, делает несколько резких поворотов на своем пути в ЦНС (рис. 4). Эти повороты происходят в тот момент, когда конус роста касается так называемых клеток-ориентиров (guidepost cells). Такое поведение указывает на наличие взаимодействия с клетками-ориентирами, ответственными за перенаправление конусов роста. Этими клетками чаше всего являются незрелые нейроны. Эти взаимодействия можно продемонстрировать при помощи удаления клеток-ориентиров лучом лазера до того, как их достигнет конус роста. В этом случае не происходит соответствующего изменения в траектории движения конуса роста.
Синаптические взаимодействия с клетками-ориентирами
В некоторых случаях аксоны образуют кратковременные синаптические контакты с клетками-ориентирами во время развития. В развивающемся гиппокампе, например, аксоны из энторинальной коры сначала образуют синапсы с временной популяцией нейронов, клетками Кахаля—Ретциуса. Позднее, по мере появления гранулярных клеток и их созревания, энторинальные аксоны покидают клетки Кахаля—Ретциуса и образуют связи с дендритами гранулярных клеток. После этого клетки Кахаля—Ретциуса исчезают. В другом примере аксоны нейронов из ядра ЛКТ в зрительной системе млекопитающих достигают развивающуюся корковую пластинку раньше, чем образуются их синаптические мишени — пирамидные клетки слоя 4. Поэтому аксоны ядра ЛКТ образуют синапсы с нейронами подпластинки , которые образуются в раннем эмбриогенезе. Нейроны подпластинки лежат под развивающейся корковой пластинкой, и им суждено исчезнуть вскоре после рождения. Через нескольких недель, когда пирамидные клетки слоя 4 достигают своего месторасположения в коре, аксоны из ЛКТ разрывают свои связи с нейронами подпластинки и направляются в кору, чтобы образовать связи, характерные для взрослого животного. Если нейроны подпластинки удалить в раннем периоде развития местной аппликацией нейротоксинов, аксоны ядра ЛКТ прорастают за пределы развивающейся зрительной коры и не могут образовать синаптических связей со своими мишенями.
Механизмы управления аксоном
Молекулы, которые управляют конусом роста, действуют четырьмя основными путями: как аттрактанты или репелленты, на короткой или длинной дистанции.
Некоторые коротко-дистантные сигналы управления аксоном обеспечиваются посредством контакта конуса роста с клеточной поверхностью или с белками адгезии внеклеточного матрикса, которые были описаны ранее в этой главе как промоторы роста аксона. Молекулы адгезии и их рецепторы могут также играть ведущую роль в связывании отдельных аксонов в пучки (так называемая фасцикуляция). Характер роста аксона и иннервация клетки-мишени in vivo и в культуре клеток нарушалась при изменении активности специфических молекул адгезии или их рецепторов, в генетических экспериментах с выключением определенных генов, а также при добавлении определенных типов антител.
Другое влияние молекул внеклеточного матрикса на растущий аксон показано в экспериментах на отдельных клетках, выделенных из ЦНС пиявки и растущих в культуре. Субстраты, которые содержат тенасцин или ламиннн, способствуют не только быстрому образованию нервных отростков у нейронов пиявки, но также влияют на характер роста этих отростков и на распределение кальциевых каналов в клетке. Различные нейроны по-разному отвечают на определенные молекулы внеклеточного матрикса. Таким образом, возникает экономная схема, когда несколько молекул адгезии могут обеспечивать разнообразные эффекты.