Регуляция дыхания
Уменьшение содержания углекислого газа в альвеолярном воздухе (и, следовательно, уменьшение напряжения его в крови) понижает деятельность дыхательного центра. Это происходит, например, в результате искусственной гипервентиляции, т. е. усиленного глубокого и частого дыхания, которое приводит к снижению парциального давления СО2 в альвеолярном воздухе и напряжения СО2 в крови. В результате наступает остановка дыхания. Пользуясь таким способом, т. е. производя предварительную гипервентиляцию, можно значительно увеличить время произвольной задержки дыхания. Так поступают ныряльщики, когда им нужно провести под водой 2…3 минуты (обычная длительность произвольной задержки дыхания составляет 40…60 секунд).
Прямое возбуждающее действие углекислоты на дыхательный центр доказано путем различных экспериментов. Инъекция 0,01 мл раствора, содержащего углекислоту или ее соль, в определенный участок продолговатого мозга вызывает усиление дыхательных движений. Эйлер подвергал изолированный продолговатый мозг кошки действию углекислого газа и наблюдал, что это вызывает увеличение частоты электрических разрядов (потенциалов действия), свидетельствующее о возбуждении дыхательного центра.
На дыхательный центр оказывает влияние повышение концентрации водородных ионов. Винтерштейн в 1911 г. высказал точку зрения, что возбуждение дыхательного центра вызывает не сама угольная кислота, а, повышение концентрации водородных ионов вследствие увеличения ее содержания в клетках дыхательного центра. Это мнение основывается на том, что усиление дыхательных движений наблюдается при введении в артерии, питающие мозг, не только угольной кислоты, но и других кислот, например молочной. Возникающая при увеличении концентрации водородных ионов в крови и тканях гипервентиляция способствует выделению из организма части содержащейся в крови углекислоты и тем самым приводит к уменьшению концентрации водородных ионов. Согласно этим экспериментам, дыхательный центр является регулятором постоянства не только напряжения углекислоты в крови, но и концентрации водородных ионов.
Установленные Винтерштейном факты нашли подтверждение в экспериментальных исследованиях. Вместе с тем ряд физиологов настаивал на том, что угольная кислота является специфическим раздражителем дыхательного центра и оказывает на него более сильное возбуждающее действие, чем другие кислоты. Причиной этого оказалось то, что углекислый газ легче, чем Н+-ион, проникает через гематоэнцефалический барьер, отделяющий кровь от цереброспинальной жидкости, которая является непосредственной средой, омывающей нервные клетки, и легче проходит через мембрану самих нервных клеток. При поступлении СО2 внутрь клетки образуется Н2СО3, которая диссоциирует с освобождением Н+-ионов. Последние и являются возбудителями клеток дыхательного центра.
Другой причиной более сильного по сравнению с другими кислотами действия Н2СО3 является, по мнению ряда исследователей, то, что она специфически влияет на некоторые биохимические процессы в клетке.
Стимулирующее влияние углекислого газа на дыхательный центр является основанием одного мероприятия, нашедшего применение в клинической практике. При ослаблении функции дыхательного центра и возникающем при этом недостаточном снабжении организма кислородом больного заставляют дышать через маску смесью кислорода с 6% углекислого газа. Такая газовая смесь носит название карбогена.
Механизм действия повышенного напряжения СО2 и увеличенной концентрации Н+-ионов в крови на дыхание. Долгое время считалось, что повышение напряжения углекислого газа и увеличение концентрации Н+-ионов в крови и цереброспинальной жидкости (ликворе) влияют непосредственно на инспираторные нейроны дыхательного центра. В настоящее же время установлено, что изменения напряжения СО2 и концентрации Н+-ионов действуют на дыхание, возбуждая находящиеся вблизи дыхательного центра хеморецепторы, чувствительные к указанным выше изменениям. Эти хеморецепторы находятся в тельцах диаметром около 2 мм, расположенных симметрично с двух сторон продолговатого мозга на вентролатеральной его поверхности поблизости от места выхода подъязычного нерва.
Значение хеморецепторов продолговатого мозга видно из следующих фактов. При воздействии на эти хеморецепторы углекислого газа или растворов с повышенной концентрацией Н+-ионов наблюдается стимуляция дыхания. Охлаждение одного из хеморецепторных телец продолговатого мозга влечет за собой, согласно опытам Лешке, прекращение дыхательных движений на противоположной стороне тела. Если хеморецепторные тельца разрушены или отравлены новокаином, дыхание прекращается.
Наряду с хеморецепторами продолговатого мозга в регуляции дыхания важная роль принадлежит хеморецепторам, находящимся в каротидном и аортальном тельцах. Это было доказано Геймансом в методически сложных опытах, в которых сосуды двух животных соединялись так, что каротидный синус и каротидное тельце или дуга аорты и аортальное тельце одного животного снабжались кровью другого животного. Оказалось, что увеличение концентрации Н+-ионов в крови и повышение напряжения СО2 вызывают возбуждение каротидных и аортальных хеморецепторов и рефлекторное усиление дыхательных движений.
Имеются данные, что 35% эффекта, вызываемого вдыханием воздуха с высоким содержанием углекислого газа, обусловлены влиянием на хеморецепторы увеличенной концентрации Н+-ионов в крови, а 65% являются результатом повышения напряжения СО2. Действие СО2 объясняется быстрой диффузией углекислого газа через мембрану хеморецептора и сдвигом концентрации Н+-ионов внутри клетки.
Рассмотрим влияние недостатка кислорода на дыхание. Возбуждение инспираторных нейронов дыхательного центра возникает не только при повышении напряжения углекислого газа в крови, но и при понижении напряжения кислорода.
Пониженное напряжение кислорода в крови вызывает рефлекторное усиление дыхательных движений, действуя на хеморецепторы сосудистых рефлексогенных зон. Прямое доказательство того, что понижение напряжения кислорода в крови возбуждает хеморецепторы каротидного тельца, получено Геймансом, Нилом и другими физиологами путем регистрации биоэлектрических потенциалов в синокаротидном нерве. Перфузия каротидного синуса кровью с пониженным напряжением кислорода приводит к учащению потенциалов действия в этом нерве (рисунок 3) и к учащению дыхания. После разрушения хеморецепторов понижение напряжения кислорода в крови не вызывает изменений дыхания.
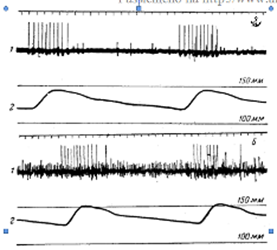
Рисунок 3 – Электрическая активность синусного нерва (по Нилу) А – при дыхании атмосферным воздухом; Б – при дыхании газовой смесью, содержащей 10% кислорода и 90% азота. 1 – запись электрической активности нерва; 2 – запись двух пульсовых колебаний артериального давления. Калибровочные линии соответствуют величине давления 100 и 150 мм рт. ст.
Запись электрических потенциалов Б показывает непрерывную частую импульсадию, возникающую при раздражении хеморецепторов недостатком кислорода. Высокоамплитудные потенциалы в периоды пульсовых повышений артериального давления обусловлены импульсацией прессорецепторов каротидного синуса.