Страница
3
В геноме бактериофагов присутствуют три типа генов: ранние, средние и поздние, классифицируемые на основании порядка их транскрипции в ходе развития фага. Ранние гены всегда транскрибируются РНКП клетки-хозяина, а в случае транскрипции средних и поздних генов возможно несколько вариантов:
1) среди продуктов ранних генов присутствует собственная фаговая РНКП, а также белки-ингибиторы РНКП хозяина. Таким образом, происходит смена РНКП;
2) среди продуктов ранних генов присутствуют новые а-факторы, взаимодействующие с «сердцевиной» РНКП хозяина и обеспечивающие транскрипцию средних генов. Среди продуктов средних генов, в свою очередь, присутствуют белки с функциями а-факторов, которые обеспечивают транскрипцию поздних генов, Таким образом, происходит смена о-факторов;
3) среди продуктов «ранних» генов присутствуют белки, модифицирующие «сердцевину» РНКП хозяина, тогда как а-фактор хозяина сохраняется.
Регуляция транскрипции путем образования специфических а-факторов широко распространена и в бактериальных системах.
Особый а-фактор контролирует транскрипцию ряда генов азотного метаболизма. Наконец, существуют промоторы, активирующиеся только при 50°, в их транскрипции участвует фактор аЕ.
Примером системной регуляции является и регуляция процесса спорообразования у бацилл. В этот процесс вовлекаются сотни локусов, организованные в специальные структуры и разбросанные по всей молекуле ДНК. Одним из способов переключения роста на спорообразование служит изменение РНКП и, в первую очередь, ее о-фактора. Такая РНКП преимущественно транскрибирует «ранние» гены спорообразования, важную роль в котором играет также регуляция на уровне трансляции.
Другой пример системной регуляции у прокариот: при воздействии на микроорганизмы нагревания до супраоптимальной температуры наступает тепловой шок, вызывающий координированную индукцию «белков теплового шока». Транскрипция их локусов и образование главного фактора а осуществляется под контролем минорного фактора с32.
Этот механизм обеспечивает координацию процессов транскрипции и трансляции и подробнее будет обсуждаться при рассмотрении способов управления скоростью роста клеток.
6. Регуляция на этапе терминации транскрипции
РНКП может «узнавать» специфические последовательности ДНК, сигнализирующие об окончании транскрипции. Эта ее способность усиливается или модифицируется под влиянием особого полипептида – р-фактора, обеспечивающего нормальную терминацию.
Наряду с этим в составе ряда регулонов обнаружены так называемые аттенюаторы, т.е. участки ДНК, вызывающие преждевременную терминацию, также р-зависимую. Эти генетические элементы располагаются между оператором и первым структурным геном в регулонах, контролирующих биосинтез ряда аминокислот. Все аминокислотные регулоны, управляемые с помощью аттенюации, характеризуются обогащением участка и РНК, соответствующего расстоянию до аттенюатора кодонами той аминокислоты, которая служит отрицательным эффектором.
Регуляция путем аттенюации основана на тесном сопряжении у прокариот транскрипции и трансляции, которые, как правило, протекают одновременно. В процессе трансляции синтезируемой иРНК рибосома, экранирующая около 10 нуклеотидов, может существенно влиять на конформацию транслируемой иРНК, а та, в свою очередь, на продвижение РНКП по матрице ДНК. При формировании «критической» конформации иРНК наступает диссоциация РНКП от ДНК.
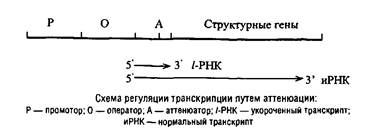
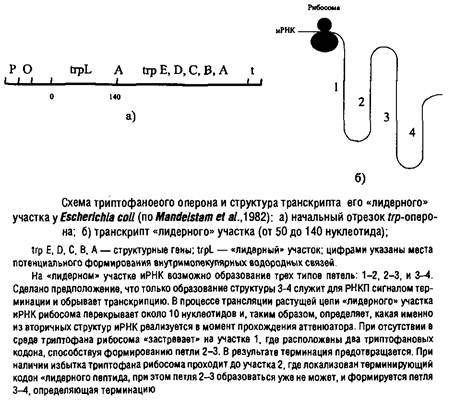
Триптофановый оперон у Escherichia coli регулируется одновременно двумя негативными механизмами: репрессией и аттенюа-цией. При избытке триптофана они действуют аддитивно, практически полностью подавляя транскрипцию. Но даже в отсутствие репрессии аттенюация снижает скорость транскрипции на 90%.
Гистидиновый оперон регулируется только посредством аттенюации. Аналогичная регуляция характерна для синтеза некоторых аминоацил-тРНК-синтетаз и, возможно, ряда экзоферментов. Аттенюатор присутствует также в геноме фага X. В этом случае эффект аттенюатора может быть нейтрализован специальным регуляторным белком антитерминатором. Получены данные о том, что комплекс белка БАК с сАМР также может выполнять функцию антитерминатора, например в опероне, и, таким образом, не только стимулировать инициацию транскрипции, но и предотвращать преждевременную терминацию.
Регуляция транскрипции путем изменения количества активной РНКП. РНКП может служить отрицательным регулятором своего собственного биосинтеза, и при относительном избытке в клетке ее дальнейшее образование тормозится. Координирование регулируется образование с переходом ее в латентное состояние. В определенных условиях латентная РНКП активируется, вызывая повышение скорости транскрипции. По некоторым данным, такая латентная РНКП может составлять до 50% всей РНКП клетки.
7. Регуляция транскрипции путем изменения конформации или структуры ДНК
Одним из способов системной регуляции транскрипции служит изменение степени сверхспирализации ДНК. В этом процессе участвует особый класс ферментов – топоизомеразы,
Релаксирующие топоизомеразы понижают степень сверхспирализации без затраты энергии и принимают участие в инициации репликации. По современной класификации их называют топоизомеразы I
Белки, повышающие степень сверхспирализации ДНК, зависят от АТР и участвуют в репликации ДНК, а также необходимы для осуществления рекомбинаций и конъюгативной передачи генетического материала. Их называют топоизомеразы II.
Косвенным указанием на возможность изменения специфичности транскрипции при изменении степени сверхспирализации ДНК служит тот факт, что в присутствии ингибиторов ДНК-гиразы изменяется спектр синтезируемых белков, хотя сами по себе эти ингибиторы не влияют на процессы транскрипции или трансляции. Например, у Escherichia coii снижается синтез около 20 белков, но одновременно стимулируется синтез других, а образование некоторых белков остается на прежнем уровне.
Перестройка транскрипции при изменении степени сверхспирализации ДНК объясняется по крайней мере двумя причинами. Во-первых, может изменяться специфичность узнавания промоторов РНК-полимеразой. Во-вторых, изменение сверхспирализованности должно приводить к изменению силы взаимодействия с ДНК регуляторных белков.
Поскольку АТР является необходимым компонентом для работы ДНК-гиразы, должна существовать связь степени сверхспирализации ДНК с энергетическим зарядом клетки, что открывает еще одну возможность для регуляции транскрипции.
Принципиально иной способ регуляции может быть осуществлен путем обратимого изменения структуры ДНК за счет выщепления подвижных генетических элементов, а затем встраивания их в другие места генома. При этом может меняться характер регуляции транскрипции как генетических локусов, входящих в состав перемещаемых участков, так и соседних с ними локусов. Возможно также встраивание генетического элемента в тот же участок ДНК, откуда он был выщеплен, но в инвертированном виде. Такая инверсия используется иногда как способ регуляции развития фагов, а также образования жгутиковых антигенов у сальмонелл.