Регенерация периферической нервной системы позвоночных
|
|
|
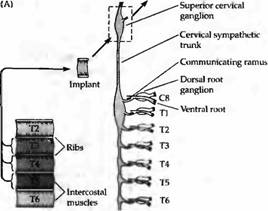
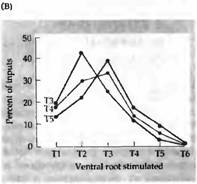
Рис. 3. Селективная реиннервация мышц, основанная на сегментарном происхождении. (А) Межреберная мышца из грудного сегмента ТЗ, Т4 или Т5 была трансплантирована на шею взрослой крысы и после удаления верхнего шейного ганглия была реиннервирована преганглионарными аксонами шейного симпатического ствола. Сегментарное происхождение входов было определено регистрацией активности мышц при стимуляции отдельных вентральных корешков, которые соединяют преганглиальные аксоны с нервным стволом. (В) Распределение входов к трансплантированным ТЗ, Т4 и Т5 мышцам. Передние мышцы склонны к реиннервации передними преганглиальными аксонами. |
Свойства нерва и мышцы после образования синапса чужим нервом
Первые наблюдения о последствиях формирования синапсов чужим нервом относятся к 1904 году, когда Лэнгли и Андерсон провели выдающееся исследование, показавшее, что мышцы кошки могут иннервироваться преганглионарными холинергическими симпатическими волокнами, которые в норме создают синапсы в автономном ганглии . Было также показано формирование подобных синапсов между автономными нервами лягушки и скелетными мышцами крысы (рис. 4.). Многие свойства нерва и мышцы в таких экспериментах оставались без изменений, несмотря на необычную иннервацию.
В других экспериментах было показано, что свойства мышц с чужеродной иннервацией заметно изменяются. Ярким примером являются медленные скелетные мышечные волокна лягушки: они иннервированы диффузно, имеют характерные детали строения и не генерируют регенеративных потенциалов действия или быстрых мышечных сокращений. После денервации медленные волокна могут быть реиннервированы нервами, которые в норме иннервируют быстрые мышцы. В этих условиях медленные волокна становятся способными генерировать потенциалы действия и осуществлять быстрое мышечное сокращение. Экклс и коллеги после перерезки меняли местами нервы, иннервирующие два типа мышц котят и крыс, различающихся по скорости сокращения. Оба этих типа мышечных волокон генерируют распространяющиеся потенциалы действия и называются медленно- и быстро-сокращающимися волокнами, соответственно. После реиннервации чужим нервом медленно-сокращающиеся мышцы становились быстрее, а быстро-сокращающиеся — медленнее. Главным фактором трансформации являлся паттерн импульсов в нерве и результирующие мышечные сокращения; мотонейроны, иннервирующие медленно- и быстро-сокращающиеся мышечные волокна, возбуждаются с различной частотой Роль базальной мембраны в регенерации нервно-мышечных синапсов
Структурой, играющей ключевую роль в регенерации нервно-мышечных синапсов, является синаптическая базальная мембрана, которая находится между нервным окончанием и мышечной мембраной. Синаптическая базальная мембрана представляет собой интенсивно окрашиваемое внеклеточное вещество, состоящее из протеогликанов и гликопротеинов. Как показано на рис. 3А, базальная мембрана окружает мышцу, нервное окончание и шванновскую клетку и погружается в складки постсинаптической мембраны.
МакМахан и его коллеги провели серию элегантных исследований роли синаптической базальной мембраны в дифференцировке нерва и мышцы. Успех их работы базировался на использовании очень удобной экспериментальной модели тонкой кожно-грудной мышцы лягушки, в которой положение концевых пластинок легко различимо в живой мышце. На первом этапе клетки в определенном участке мышцы локально разрушали либо перерезкой нерва и мышечных волокон, либо повторным приложением металлической пластины, охлажденной жидким азотом (рис. 3В). В течение нескольких дней часть мышечных волокон в участке повреждения дегенерировала вместе с нервными окончаниями и была фагоцитирована, но базальная мембрана оставалась при этом интактной (рис. 3C). Место исходного нервно-мышечного соединения могло быть легко распознано по характерной морфологии базальной мембраны мышцы и шванновских клеток, остающихся в зоне синапса, а также по наличию холинэстеразы, сохраняющейся на синаптической базальной мембране и в синаптических складках.
Через две недели после повреждения в зоне синаптической базальной мембраны формировались новые мышечные волокна, контактирующие с регенерирующими аксональными окончаниями. Стимуляция нервов вызывала сокращения вновь образованных мышечных волокон. Почти все регенерированные синапсы локализовались точно на исходных синаптических зонах, что выявлялось после окраски мышцы на присутствие холинэстеразы. Таким образом, был сделан вывод о наличии сигналов, ассоциированных с синаптической базальной мембраной, которые определяют успех регенерации синапсов.
Синаптическая базальная мембрана и формирование синаптической специализации
Для дальнейшего исследования природы сигналов, ассоциированных с синаптической базальной мембраной, мышцы повреждали, нерв раздавливали, а регенерацию мышечного волокна предотвращали рентгеновским облучением. Регенерирующие аксоны росли к исходным синаптическим зонам, что выявлялось окраской на холинэстеразу, и формировали активные зоны для освобождения медиатора точно напротив участков базальной мембраны, связанных со вторичными синаптическими складками, — и все это происходило в отсутствие клеточных элементов постсинаптической мишени (рис.3D).
В параллельной серии экспериментов МакМахан и его коллеги продемонстрировали, что синаптическая базальная мембрана в регенерирующих мышечных волокнах содержит факторы, запускающие дифференцировку постсинаптической мембраны. Мышцы повреждались, как было описано ранее, а реиннервация предотвращалась удалением большого сегмента нерва. При регенерации новые мышечные волокна образовывали вторичные складки и кластеры АХ рецепторов и ацетилхолинэстеразы точно в зоне контакта с исходной синаптической базальной мембраной (рис. 4). Таким образом, сигналы, ассоцированные с синаптической базальной мембраной, при регенерации могут инициировать формирование синаптических специализаций как в мышечных волокнах, так и в нервных окончаниях.
Идентификация агрина
Для идентификации сигнала, связанного с базальной мембраной и инициирующего постсннаптическую дифференцировку, МакМахан и его коллеги использовали морского ската Torpedo califomica. Из электрических органов этого животного, родственных скелетной мышце, они приготовили экстракты, содержащие базальные мембраны. Добавленные к культуре мышечных волокон, экстракты симулировали эффекты синаптической базальной мембраны на регенерацию мышечных волокон, а именно индуцировали формирование кластеров АХ рецепторов вместе с другими компонентами постсинаптической мембраны (рис. 5).
Активный компонент экстрактов, названный агрином, был очищен и охарактеризован, а у цыпленка, крысы и ската клонирована соответствующая кДНК.