Страница
2
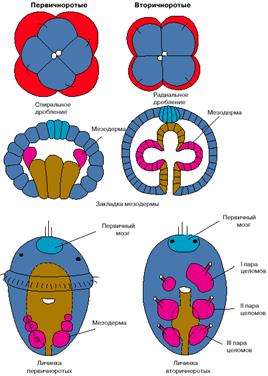
Рис. 1. Особенности эмбрионального развития первичноротых и вторичноротых животных.
Сравнение хордовых с другими вторичноротыми
Хотя хордовые - это вторичноротые животные, все же их развитие имеет ряд особенностей. На рис. 3 представлены соответствующие стадии развития одной из групп вторичноротых - погонофор и низших хордовых - асцидий.
Анимальный полюс яйца погонофор, на котором располагается ядро яйцеклетки, соответствует спинной стороне эмбриона, а вегетативный, на котором концентрируется желток, - брюшной. У низших хордовый ситуация обратная: анимальный полюс соответствует брюшной стороне, а вегетативный - спинной. Бластопор (то есть та область эмбриона, где впячивается энтодерма и формируется первичный кишечник) у погонофор соответствует брюшной стороне личинки, а у низших хордовых - спинной. Зачаток нервной системы, закладывающийся по линии замыкания бластопора, у погонофор становится брюшным нервным тяжем, а у хордовых - спинной нервной трубкой (рис. 3).

Рис. 2. Представители основных групп первичноротых и вторичноротых животных.
План строения низших хордовых также существенно отличается от планов строения других вторичноротых. Так, например, у всех вторичноротых передние целомы диссимметричны: левый передний целом больше правого, который обычно выполняет функцию околосердечной сумки. У хордовых ситуация обратная: правый передний целом больше левого. Целомы вторичноротых сообщаются с внешней средой отверстиями, открывающимися на спинной стороне и выполняющими осморегуляторные и половые функции. У хордовых эти отверстия располагаются на той стороне тела, которую мы традиционно считаем брюшной.
Кровеносная система у вторичноротых и хордовых устроена по общему плану и состоит из двух главных сосудов: спинного и брюшного. У вторичноротых, как и всех других беспозвоночных, кровь по спинному сосуду течет вперед, а по брюшному назад. У хордовых направление тока крови обратное: по спинному сосуду кровь течет назад, а по брюшному - вперед. Сердце у вторичноротых лежит на спинной стороне, а у хордовых - на брюшной.
Суммируем все эти отличия.
1. Бластопор хордовых соответствует не брюшной, а спинной стороне.
2. Нервная пластинка хордовых, закладывающаяся по месту замыкания бластопора, превращается не в брюшной нервный тяж, а в спинную нервную трубку.
3. Диссимметрия первой пары целомов имеет обратный знак в сравнении с другими вторичноротыми: правый передний целом у хордовых больше левого.
4. Отверстия целомов у хордовых открываются не на спинной, а на брюшной стороне.
5. Кровь у хордовых по брюшной стороне течет вперед, а по спинной - назад, и сердце располагается не на спинной, а на брюшной стороне.
Что же означают эти несоответствия? Только одно: предки хордовых в своем эволюционном развитии испытали инверсию сторон тела, то есть перевернулись и стали передвигаться на морфологически спинной стороне, которая стала функционировать как физиологически брюшная. Морфологически брюшная сторона предков хордовых стала функционировать как физиологически спинная.
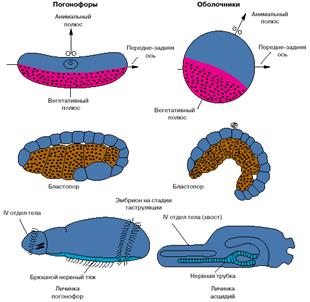
Рис. 3. Особенности эмбрионального развития погонофор и низших хордовых.
Если мы примем этот вывод, все перечисленные выше несоответствия исчезнут: бластопор хордовых станет соответствовать их исходно брюшной стороне, нервная трубка будет располагаться на морфологически брюшной стороне, диссимметрия передних целомов приобретет нормальный для вторичноротых характер, отверстия целомов окажутся на морфологически спинной стороне, и направление тока крови, так же как положение сердца, окажется типичным для вторичноротых. На рис. 4 представители низших хордовых (личинка асцидий и ланцетник) изображены перевернутыми, чтобы продемонстрировать, что при таком положении их системы органов соответствуют таковым других вторичноротых.
Переворот на спинную сторону мог стать тем важнейшим эволюционным событием, которое определило формирование плана строения хордовых животных и его коренное отличие от планов строения вторичноротых и других беспозвоночных.
Происхождение хордовых животных. В. В. МАЛАХОВ. Продолжение.
Пути эволюции вторичноротых животных
Общий предок вторичноротых животных, по-видимому, вел донный образ жизни, ползая на брюшной стороне с помощью ресничной полоски, подостланной вентральным нервным тяжем (рис. 5). Он имел три пары целомических мешков и щупальцевый аппарат, с помощью которого собирал мелкие пищевые частицы. В дальнейшем могло произойти обособление самого заднего отдела тела, с помощью которого предки вторичноротых временно зарывались в верхние слои грунта. В эволюции вторичноротых этому отделу принадлежит очень важное место.
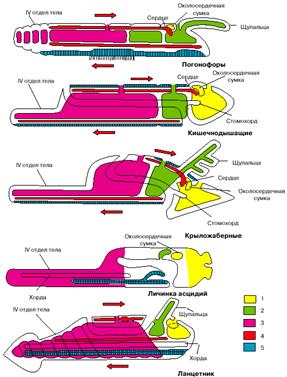
Рис. 4. Сравнение планов строения различных групп вторичноротых (личинка асцидий и ланцетник изображены в перевернутом положении). 1 - целомы первой пары, 2 - целомы второй пары, 3 - целомы третьей пары, 4 - кровеносная система, 5 - нервная система. Стрелки показывают направление тока крови.
Предки иглокожих перешли к сидячему образу жизни и стали собирать взвешенные в воде пищевые частицы с помощью щупалец. Радиальная симметрия иглокожих - это наследие сидячего образа жизни, который среди современных форм сохраняют морские лилии, тогда как морские звезды, морские ежи, офиуры и голотурии способны медленно переползать с места на место.
Полухордовые в качестве основного органа движения стали использовать предротовой отдел тела:
хобот кишечнодышащих или головной щит крыложаберных. Обычно полухордовых считают близкими предками хордовых, о чем говорит и само название этой группы. В действительности эта группа не слишком близка к хордовым, в частности потому, что имеет два нервных тяжа, только один из которых может быть гомологизирован с нервной трубкой хордовых. У них имеется слепой вырост кишечника - стомохорд, который сформировался как опорная структура хобота и не имеет прямого отношения к хорде хордовых животных.

Рис. 5. Филогенетические отношения между вторичноротыми животными.
Современные погонофоры - сидячие организмы, обитающие внутри выделяемых ими трубок. Во взрослом состоянии они лишены кишечника и питаются за счет симбиотических хемосинтезирующих бактерий, окисляющих сероводород или метан. Их предки, вероятно, питались этими бактериями, собирая их из придонного слоя воды с помощью щупалец. Впоследствии они стали культивировать бактерий в клетках кишечника. Личинки одной из групп современных погонофор - вестиментифер - на ранних стадиях развития собирают бактерий с поверхности грунта, а затем начинают их культивировать внутри видоизмененного кишечника, после чего рот и анус у них редуцируются. Особенности эмбрионального развития и строения нервной системы позволяют рассматривать погонофор как самую близкую к хордовым группу вторичноротых.