Страница
2
Кинетические уравнения как простых каталитических, так и автокаталитических систем при постоянной концентрации субстрата содержат член первого порядка по концентрации катализатора. Кривые роста, однако, четко дифференцируют две эти системы. Количество продукта для простого каталитического процесса в указанных условиях растет во времени линейно, тогда как для автокаталитических систем характерен экспоненциальный рост.
Строго говоря, автокаталитическую систему уже можно называть гиперциклической, ибо она представляет собой цикл катализаторов, которые сами являются циклами реакций. Мы, однако, ограничим использование этого термина такими системами, которые являются гиперциклическими по отношению к каталитической функции. В действительности это гиперциклы второй или более высокой степени, потому что они относятся к реакциям по меньшей мере второго порядка по концентрациям катализаторов.
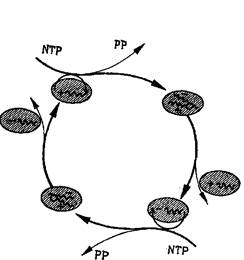
Рис. 5. Каталитический цикл, имеющий биологическое значение, — репликация одноцепочечной РНК.
Интермедиаты (плюс- и минус-цепи) участвуют в цикле как матрицы для своего взаимного воспроизведения. Матричная функция эквивалентна избирательному катализу. Нуклеозидтрифосфаты (NTP) представляют собой высокоэнергетический строительный материал, а пирофосфат (РР) является отходом реакций цикла. Комплементарное инструктирование, механизм которого будет обсуждаться в связи с рис. 11, является существенно автокаталитической функцией, т. е. функцией самовоспроизведения.

Рис. 6. Истинный процесс самовоспроизведения (одночленный каталитический цикл) — репликация ДНК.
Механизм этого чрезвычайно сложного процесса (см. рис. 12) обеспечивает связь каждой дочерней цепи (D) с соответствующей родительской цепью (Р).

Рис. 7. Каталитический гиперцикл состоит из самоинструктируюшихся единиц U с двойными каталитическими функциями.
В качестве автокатализатора или — в более общем случае—в качестве каталитического цикла ннтермедиат U способен инструктировать свое собственное воспроизведение, а кроме того, он оказывает каталитическое содействие воспроизведению следующего интермедиата (с использованием высокоэнергетичеекого строительного материала X). Упрощенная схема (Б) показывает иерархию циклов.
Каталитический гиперцикл — это система, связывающая автокаталитические, или самовоспроизводящиеся единицы друг с другом посредством циклической связи. Такая система изображена на рис. 7. как самовоспроизводящиеся единицы сами являются каталитическими циклами, например комбинациями плюс- и минус-цепей РНК, как показано на рис. 5. Однако процесс репликации как таковой должен прямо или косвенно стимулироваться дополнительными специфическими связями между различными репликативными единицами.
Реально такие связи могут обеспечиваться белками — продуктами трансляции предшествующих циклов воспроизведения РНК (рис. 8). Эти белки могут действовать как специфические репликазы или дерепрессоры или же как специфические защитные факторы, предохраняющие от деградации. Связи между самовоспроизводящимися единицами должны образовывать наложенный цикл — лишь тогда система в целом будет подобна гиперциклу. По сравнению с системами, показанными на рис. 4 и 5, гиперцикл является самовоспроизводящимся в более высокой степени.
Простейшим представителем систем этого класса снова является (квази)одношаговая система, т.е. усиленный автокатализатор. Мы сталкиваемся с такой системой в случае инфекции клетки РНК.

Рис. 8. Реалистическая модель гиперцикла второй степени, где носители информации «выдают» инструкции двух типов: во-первых, для собственного воспроизведении и, во-вторых, для трансляции с образованием интермедиатов второго типа, которые обладают оптимальными функциональными свойствами.
Каждый фермент Е; катализирует воспроизведение следующего носителя информации I;. Он может иметь и другие каталитические свойства, необходимые для осуществления процесса трансляции, метаболизма и т.д. В подобных случаях степень гиперциклической связи больше двух.
Если фаговая РНК (плюс-цепь) проникает в бактериальную клетку, то ее генотипическая информация транслируется аппаратом клетки-хозяина. Один из продуктов трансляции связывается с определенными факторами хозяина с образованием активного ферментного комплекса, который специфически реплицирует плюс- и минус-цепи фаговой РНК, действующие при воспроизведении как матрицы друг для друга [11]. Однако репликазный комплекс не размножает — в сколько-нибудь ощутимой степени — информационную РНК клетки-хозяина. В результате инфекции начинается гиперболический рост числа фаговых частиц, который в конце концов выходит на насыщение из-за ограниченности ресурсов клетки-хозяина.
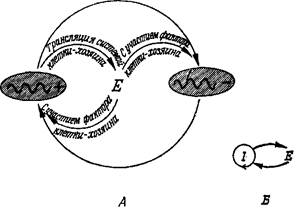
Рис. 9. Размножение РНК-содержащего фага в бактериальной клетке представляет собой простой гиперциклнческий процесс.
Инфекционная плюс-цепь, используя аппарат трансляции клетки-хозяина, сначала инструктирует синтез белковой субъединицы (Е), которая связывается с другими белками клетки-хозяииа с образованием фагоспецифичной РНК-репликазы. Этот репликазный комплекс узнает только определенные фенотипические особенности фаговой РНК, которые имеются как у плюс-, так и у минус-цепи благодаря симметрии определенных участков цепи РНК. В результате происходит вспышка синтеза фаговой РНК, которая — из-за гиперциклической природы — носит гиперболический характер (см. часть Б, рис. 17), пока не наступит насыщение по одному из интермедиатов или пока не истощатся запасы метаболитов клетки-хозяина. Схема Б показывает, что автокаталитическими илк самоинструктивиыми свойствами может обладать всего один из интермедиатов при условии, что другие «партнеры» связаны с ним с помощью циклической обратной связи.

Рис. 10. Иерархия циклических реакционных сетей очевидна из сопоставления этих схем (— химическое превращение, —* каталитическое действие).
Другой естественный гиперцикл может возникнуть в менделеевских популяциях в начальной фазе видообразования — пока численности популяций малы. Для репродукции генов необходимо взаимодействие между двумя аллелями (М и F), т. е. между гомологичными участками в мужской и женской хромосомах, которые затем появляются в потомстве в перестроенной комбинации. Тот факт, что кинетические уравнения популяционной генетики менделеевских популяций обычно не содержат гиперциклической нелинейности (которая ведет к гиперболическому, а не к экспоненциальному росту), объясняется насыщением, наступающим при сравнительно малых численностях популяций, когда скорость рождений (обычно) становится пропорциональной только численности самок.
Как видно из сравнения схем, показанных на рис. 10, гиперциклы представляют собой новый уровень организации. Этот факт проявляется в их уникальных свойствах. Не связанные друг с другом самовоспроизводящиеся единицы гарантируют сохранение ограниченного количества информации, которое может передаваться от поколения к поколению. Это оказывается одной из необходимых предпосылок дарвиновского поведения, т. е. отбора и эволюции. Подобно им, гиперциклы тоже селекционны, но, кроме того, они имеют интегрирующие свойства, которые делают возможной кооперацию между единицами, конкурирующими в других условиях. Более того, гиперциклы конкурируют даже еще более ожесточенно, чем дарвиновские виды, с любой самовоспроизводящейся единицей, не являющейся частью гиперцикла. Далее, они способны устанавливать глобальные формы организации благодаря свойству отбирать «раз и навсегда», которое не допускает сосуществования с другими гиперциклическими системами, если только они не стабилизированы связями более высокого порядка.