Передача и кодирование сигнала в сетчатке глаза
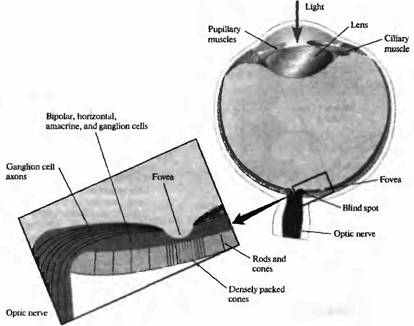
Рис. 3. Путь света в полости глаза и клеточное строение сетчатки (поперечный срез глаза). Свет должен пройти через хрусталик и слои клеток сетчатки, чтобы достигнуть фоторецепторов: палочек и колбочек. Центральная ямка (fovea centralis) является специализированной зоной, содержащей только густо расположенные тонкие колбочки, которые способны различать объекты с высоким разрешением. В этом месте слои клеток, расположенные более поверхностно, менее выражены, что позволяет свету непосредственно падать на фоторецепторы. В месте, где зрительный нерв выходит из глаза, отсутствуют фоторецепторы, и потому это место называется "слепым пятном".


Рис.4. Фоторецепторы сетчатки. (А и В) Палочка сетчатки жабы с введенным в нее флуоресцентным красителем, Lucifer yellow, при обычном (А) и ультрафиолетовом (В) облучении. Стрелками отмечены одинаковые места в сетчатке. (С) Строение палочки и колбочки. В палочке пигмент родопсин (черные точки) встроен в мембраны, расположенные в виде дисков, не связанных с наружной мембраной клетки. В колбочке зрительные пигменты располагаются в складках наружной клеточной мембраны. Наружный сегмент связан с внутренним посредством узкой перемычки. В темноте происходит постоянное высвобождение медиатора нервными окончаниями.
При помощи различных подтипов колбочек мы способны воспринимать мельчайшие оттенки и изменения в контрастности и цвете в яркий солнечный день, когда интенсивность света в 100 миллионов раз сильнее, чем ночью.
Организация и морфология фоторецепторов
Палочки и колбочки образуют плотно организованный слой фоторецепторов, расположенный непосредственно над слоем пигментного эпителия (рис.3), на максимальном удалении от роговицы и проникающего в глаз света. За исключением небольшой зоны в области центральной ямки, свет должен пройти через несколько плотных слоев клеток и волокон, чтобы достигнуть наружного сегмента фоторецептора, где и осуществляется поглощение фотонов и фоторецепция. Как Гельмголъц писал в 1867открыт синапс и вообще появилась клеточная доктрина*).
Исследования центральной ямки показали, что в этой зоне колбочки очень плотно упакованы, их плотность составляет 200000 на мм2, а палочки отсутствуют. Более того, в области ямки колбочки более узкие, чем в периферических зонах сетчатки. Так как ямка не содержит палочек, ночью она представляет собой "слепое пятно". Существует и отдельное "слепое пятно", которое соответствует зоне на сетчатке, где зрительный нерв покидает глаз; в зоне этого пятна отсутствуют любые фоторецепторы.
На рис.4 показаны три важные особенности строения фоторецепторов: (1) наружный сегмент, где происходит поглощение света зрительными пигментами; (2) внутренний сегмент, где расположены ядро, ионные насосы, трансмембранные переносчики, рибосомы, митохондрии и эндоплазматический ретикулум; и (3) синаптическая терминаль, где происходит высвобождение глутамата на клетки второго порядка, чувствительные к глутаматному сигналу. Зоны секреции в синаптической терминали имеют строго определенное расположение в зависимости от расположения одной или нескольких "ленточных" структур, вдоль которых ориентированы везикулы с медиатором.
Хотя сама по себе "клеточная доктрина", или "клеточная теория", была опубликована Маттиасом Шлейде ном и Теодором Шванном в 1839 году ("Микроскопические исследования о соответствии в структуре и росте животных и растений"), она содержала несколько ошибочных утверждений (например, о новообразовании клеток организма из первичного неклеточного вещества), и получила известность и распространение значительно позже, после уточнения ее Рудольфом Биржевым в 1859 году
Электрические сигналы в ответ на свет в фоторецепторах позвоночных
Рецепторы в основном отвечают на соответствующий стимул путем локальной деполяризации переменной амплитуды, которая может вызвать в дальнейшем потенциал действия. И хотя у большинства беспозвоночных фоторецепторы воспринимают свет подобным образом (рис.5А), восприятие света рецепторными клетками у большинства позвоночных происходит по-другому. На рис.5В показаны сигналы, отведенные от палочки черепахи при помощи внутриклеточного электрода. В темноте (во время отдыха) фоторецепторы деполяризуются постоянным входящим током в области наружного сегмента ("темновой ток"). Свет приводит к гиперполяризации, уменьшая величину входящего тока. В следующих абзацах будет рассказано, каким образом свет поглощается фоторецепторами и какие механизмы вызывают электрические сигналы, показанные на рис. 5.
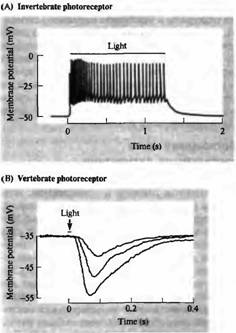
Рис. 5. Сигналы от фоторецепторов. (А) Фоторецепторы беспозвоночных (мечехвост) отвечают на свет деполяризацией, вызывающей серию разрядов. Такие ответы являются типичными для сенсорных клеток, чувствительных к различным стимулам: прикосновению, давлению или растяжению (глава 17). (В) Фоторецепторы позвоночных (черепахи) отвечают на свет гиперполяризацией, уровень которой зависит от интенсивности вспышки.
Слуховая кора
Слуховой вход, осуществляемый как через дорзальные, так и вентральные кохлеарные ядра, достигает слуховой коры. Первичная слуховая кора (А,) расположена в верхнем отделе височной доли и соответствует полям 41 и 42 по Бродману. У кошек А, удобно расположена на боковой поверхности мозга, поэтому большинство комбинированных анатомо-физиологических исследований производится на этом виде. Микроэлектродные исследования показали, что А, также имеет колончатую организацию, при которой все клетки, расположенные вдоль вертикального пути, имеют одну и ту же оптимальную частоту. Фактически A1 организована в виде изочастотных полос или пластин коры, идущих перпендикулярно к тонотопической оси.
Слуховая кора у обезьян содержит три полных кохлеотопических карты с параллельными проекциями из ядер медиального коленчатого тела на все эти области. Самая задняя из них соответствует первичной слуховой коре, как показано на основе ее гистологических свойств. Эта центральная основа окружена вторичными слуховыми областями, которые соединены не только с первичной корой, но также и с подразделами ядер медиального коленчатого тела. Таким образом, в слуховой коре происходит как последовательная, так и параллельная обработка. Исследования с помощью электроэнцефалографического картирования и функционального магнитного резонанса представляют доказательства подобной же организации слуховой коры у человека, и эта область коры активирована даже во время чтения по губам!)
По аналогии с другими сенсорными участками коры можно ожидать, что кохлеотопическая карта А, будет подразделена на различные функциональные зоны. Если двигать электрод вдоль изочастотной полоски, большинство клеток либо возбуждаются звуком, воспринимаемым любым из ушей (ЕЕ), либо возбуждаются из одного уха и тормозятся из другого (EI). Эта бинауральная чувствительность закономерно меняется, и полагают, что ее значения образуют полосы, идущие под прямым углом к изочастотным контурам. Существуют также некоторые доказательства закономерных изменений кодирования интенсивности и ширины полосы в A1. Организация ЕЕ-EI может быть выведена из бинауральных взаимодействий, которые сначала возникают в ядрах олив, и приводит к предположению, что А, также картирована для слухового пространства, хотя в настоящее время это не доказано.