Страница
2
У рода вошерия (Vaucheria) (известны как пресноводные, так и морские и солоноводные виды) таллом в виде разветвленных сифоновых нитей, образующих ватообразные скопления в воде или обширные дерновинки темно-зеленого цвета на влажной земле. Рост нитей апикальный. В вегетативных нитях вошерии, начиная с верхушки, можно различить три зоны: апикальную, субапикальную и вакуолизированпую. Апикальный отрезок представляет собой активно растущую часть сифонового таллома: здесь находятся многочисленные пузырьки и митохондрии; хлоропласты и клеточные ядра отсутствуют. Везикулы содержат фиброзный материал, возможно, являющийся предшественником материала (целлюлозы) клеточной оболочки. В субапикальной зоне количество пузырьков уменьшается, появляются хлоропласты и клеточные ядра. Хлоропласты многочисленные, дисковидные, лишены пиреноидов. Выдающийся в виде почки пиреноид наблюдается только в проростках вошерии. С каждым клеточным ядром связана пара центриолей, которые во время профазы митоза мигрируют к противоположным полюсам удлиняющихся ядер. Оболочка ядра во время митоза остается интактной, внутри нее образуется внутриядерное веретено, центромеры отсутствуют; между расходящимися хромосомами ядерная оболочка перешнуровывается и окружает телефазные дочерние ядра. Нормальный митоз, происходящий внутри оболочки материнского ядра, у ряда видов вошерии наблюдал еще Л. И. Курсанов (1911). У всех исследованных видов Л.И. Курсанов отметил интересное распределение ядерных делений на протяжении нити: начавшись в одном каком-нибудь месте, митозы мало-помалу распространяются на соседние участки и т. д. Таким образом, на препарате, фиксирующем известный момент этого процесса, получается то, что всего лучше назвать волной делений, где отдельные, следующие друг за другом во времени фазы деления на протяжении нити расположены в правильной последовательности. В апикальной и субапикальной зонах еще отсутствует центральная вакуоль. Лишь в более старой — вакуолизированной — части нити появляется вакуоль с клеточным соком (рис. 23, А). Размножение бесполое, посредством синзооспор и апланоспор.
Образование синзооспор, их выход, оседание и прорастание были прослежены у Vaucheria fontinalis па электронно-микроскопическом уровне (рис. 23,5"—Е). При образовании синзооспоры на конце нити, несколько вздутом, центральная вакуоль исчезает, и здесь скапливаются все органеллы. Связанные с клеточными ядрами пары центриолей формируют внутренние жгутики. Клеточные ядра и внутренние жгутики группируются вокруг возникающих пузырьков, в которые вдаются жгутики. Пузырьки мигрируют к поверхности протопласта и сливаются с плазмалеммой. Таким образом, жгутики достигают поверхности будущей синзооспоры.
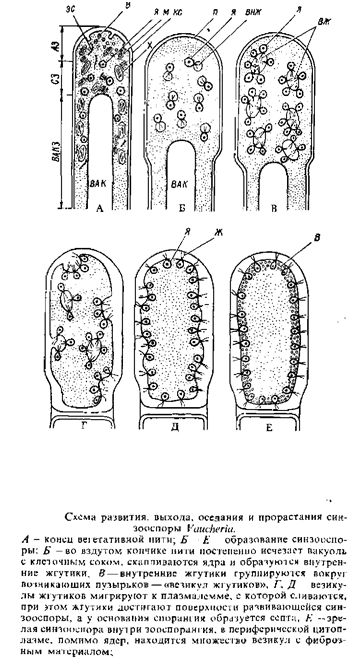
Синзооспоры вошерии покрыты многочисленными гладкими, очень незначительно различающимися по длине парами жгутиков. Возникает септа, отделяющая спорангий с синзооспорой от вегетативной нити. Выход синзооспоры, по-видимому, осуществляется после энзиматического растворения верхушки зооспорангия. Недавно остановившаяся синзооспора сферическая, крупная, центральная вакуоль отсутствует, жгутики втягиваются, сохраняя сначала типичную (9 — 9 + 2) конфигурацию аксонемы; их мембраны становятся частью плазмалеммы. ядра занимают субповерхностную зону. Клеточной стенки сначала нет, но в периферическом слое цитоплазмы находятся обильные везикулы с фибриллярным материалом, которые, по-видимому, содержат предшественников целлюлозы. Их исчезновение совпадает с отложением на поверхности синзооспоры тонкой стенки. Ядра смещаются, в периферическую область вторгаются хлоропласты и другие органеллы. Аксонемы втянутых жгутиков разрушаются, центриоли занимают свое обычное положение рядом с клеточными ядрами. Образуется крупная центральная вакуоль. С одного или с обоих концов инцистированной зооспоры возникает выпуклость, в которой скапливаются везикулы с фибриллярным материалом, митохондрии, диктиосомы, хлоропласты, ядра, как это наблюдается в кончике вегетативной нити. Центральная вакуоль инцистированной зооспоры начинает удлиняться (рис. 23,Ж—Л),
Половой процесс оогамный. Пресноводные виды вошерии большей частью однодомные, морские виды-двудомные.
Половые органы возникают в виде боковых выростов нити, содержащих многочисленные ядра: более или менее шаровидной формы в случае оогония и в форме цилиндрической трубки, закрученной наподобие рога - при развитии антеридия. Оба типа половых выростов сначала сообщаются с вегетативной нитью, на которой заложились, а позднее отделяются от нее поперечной стенкой. В случае оогония до образования поперечной перегородки все ядра, кроме одного, становящегося ядром единственной яйцеклетки, мигрируют с очень подвижной цитоплазмой обратно в вегетативную нить. По другим данным, лишние ядра дегенерируют после отделения оогония от нити поперечной перегородкой. В антеридии, который отделяется поперечной перегородкой от кончика мужского выроста, сохраняются многочисленные ядра и возникает большое число сперматозоидов. Сперматозоид вошерии без хлоропласта и стигмы, с крупными митохондриями, снабжен двумя гетероконтными жгутиками, причем передний жгутик с мастигонемами короче гладкого заднего. В передней части сперматозоида имеется вырост с микротрубчатыми скелетными элементами, напоминающий так называемый хоботок сперматозоида фукуса. Половые органы вскрываются, сперматозоиды проникают в оогоний и оплодотворяют яйцеклетку, которая после оплодотворения выделяет толстую оболочку. Копуляционное ядро в прорастающей после периода покоя ооспоре редукционно делится.
Некоторые исследователи, придавая большое значение различиям в строении жгутикового аппарата у сперматозоидов и синзоосиор вошерии, выделили этот род даже в отдел Vaucheriophyta, поместив остальные желтозеленые водоросли в качестве класса Heteroconlae в отдел Chrysophyta. Другие авторы объединяли роды ботридиум и вошерия в отдел Siphonophyta, позднее понизив этот таксон до ранга класса Xanthosiphonophyceae наряду с Xanthophyceae в пределах отдела Chrysophyta. Сегрегация рода вошерия от других желтозеленых водорослей на столь высоком уровне, по-видимому, не встретила поддержки у многих исследователей. Однако выделение вошерии в отдельный порядок Vaucheriales принято многими авторами.
Класс эвстигматофициевые (Eustigmatophyceae)
Этот класс был выделен из ксантофициевых на основе строения монадных клеток (зооспор), в первую очередь их глазка. Глазок представляет собой крупное оранжево-красное тело у крайнего переднего конца зооспоры, не зависимое от единственной пластиды, и состоит из неправильной группы капель без ограничивающих мембран и без мембраны вокруг всего комплекса. Таким образом, в отличие от ксантофициевых стигма здесь расположена вне пластиды. Другой отличительный признак эветигматофициевых — то, что утолщение жгута, перекрывающее стигму, находится у проксимального конца, направленного вперед и несущего два ряда волосков жгутика. Гладкий, направленный назад жгутик, как правило, отсутствует. Кроме того, у эвстигматофициевых трехтилакоидные ламеллы часто образуют граноподобные стопки, периферическая опоясывающая ламелла отсутствует, пиреноид, обнаруженный только в вегетативных клетках (в зооспорах отсутствует), выступает от внутренней поверхности пластиды и не пересечен тилакоидами.