Страница
4
В рассмотренной схеме также наблюдалось взаимодействие генов в F2, в результате которого дрозофилы имели нормальный цвет глаз. Такой тип взаимодействия носит название комплементарности или комплементарного (взаимно дополнительного) действия, когда доминантные аллели обоих генов обусловили нормальный (или дикий) фенотип (под комплементарностью обычно подразумевают именно этот тип взаимодействия генов).
В F2, рецессивные аллели тех же генов обусловили появление белоглазых мух. Был приведен пример того, что носит название формально-генетического анализа, при котором полностью абстрагируются от механизмов действия исследуемых генов. Если анализ проведен верно и формальные отношения генов и аллелей выявлены правильно, то последующее выяснение физиологических механизмов, лежащих в основе генных взаимодействий, только подтверждает и конкретизирует выводы.
Биохимический механизм взаимодействия аллелей генов st и bw исследован достаточно подробно. Известно, что у дрозофилы окраска глаз обусловлена синтезом двух пигментов - красного и бурого. Рецессивная аллель bw в гомозиготе прерывает синтез красного пигмента, поэтому глаза содержат только бурый пигмент. Рецессивная аллель st в гомозиготе блокирует синтез бурого пигмента, вследствие чего в глазах мух содержится только красный пигмент.
Когда в дигетерозиготе оказываются нормальные аллели обоих генов, синтезируются оба пигмента. Результат - комплементарное взаимодействие нормальных аллелей, наблюдаемое в F1. Если в F2 в гомозиготе оказываются и bwbw, и stst, то не синтезируются ни красный, ни бурый пигменты, и глаза оказываются белыми, неокрашенными.
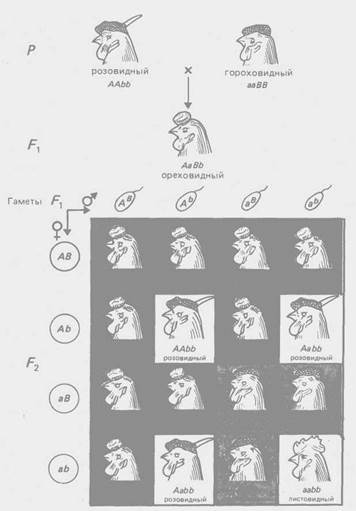
Рис. 2 - Комплементарное взаимодействие генов, определяющих форму гребня у кур (соотношение 9:3:3:1)
Можно рассмотреть и другой тип комплементарного взаимодействия генов у дрозофилы, если идти от метаболического эффекта генов к схеме их взаимодействия. Вновь обратимся к биосинтезу пигментов глаза у дорозофилы (рис. 3). Известно, что кроме рецессивных аллелей st синтез бурого пигмента блокируют и рецессивные аллели гена purple (pr). Фенотип гомозигот stst и ргрт - ярко-красные глаза. При скрещивании таких мух в F1 глаза нормальные - темно-красные, поскольку работают оба гена - комплементарно взаимодействуют их доминантные аллели. В F2 наблюдается следующее соотношение фенотипов: 9 с темно-красными и 7 с ярко-красными глазами. Это объясняется тем, что выход в гомозиготу любой из двух рецессивных аллелей достаточен для блокирования синтеза пигмента, тем более, когда и рг, и st находятся в гомозиготе. Это тоже пример комплементарного взаимодействия, но без новообразования в F2.
По типу комплементарное™ взаимодействуют гены, контролирующие разные этапы одного и того же метаболического пути. Однако для многих морфологических признаков неизвестен биохимический механизм их реализации, поэтому приходится ограничиваться констатацией формально-генетической схемы их наследования. Так, по типу комплементарности взаимодействуют гены, определяющие форму гребня кур (рис. 2), форму плода у тыквы (рис. 3) и др.
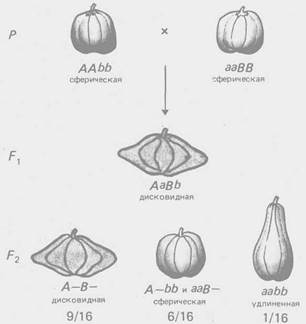
Рис. 3 - Комплементарное взаимодействие генов, определяющих форму плода тыквы (соотношение 9:6: 1)
3.2 Эпистаз
Вернемся к анализу взаимодействия генов рr и st у дрозофилы. Соотношение фенотипических классов в F2 можно представить себе и как следствие того, что рецессивная аллель рr в гомозиготе препятствует проявлению доминантной аллели st+. Точно так же рецессивная аллель st в гомозиготе препятствует проявлению доминантной аллели рr+. Действительно, то, что известно о генетическом контроле синтеза бурого глазного пигмента у дрозофилы, вполне соответствует предложенному здесь объяснению. Такой тип взаимодействия носит название эпистатического, или эпистаза, и условно изображается: рr > st+ и st > pr+. В данном случае рецессивная аллель рr эпистатична по отношению к доминантной аллели st+, a st эпистатична по отношению к рr+. Данный случай взаимодействия генов называют также двойным рецессивным эпистазом.
По изменению числа и соотношения классов дигибридного расщепления в F2 рассматривают несколько типов эпистатических взаимодействий: простой рецессивный эпистаз (а > В; а> Ь или b > А; Ь > а), который выражается в расщеплении 9:3:4; простой доминаданый эпистаз (А> В; А> b или В > А; B > а) с расщеплением 12:3:1 и т.д.
Один ген, подавляющий действие другого, называют эпистатическим геном, ингибитором или супрессором. Подавляемый ген носит название гипостатического.
Как уже показано, констатация того или иного типа взаимодействия генов в дигибридном скрещивании условна. Тем не менее, при кажущемся нарушении закона независимого наследования (появлении неожиданных классов в расщеплении или уменьшении числа классов), связанного с взаимодействием двух генов, всегда можно свести наблюдаемые соотношения в F2 к классическому 9:3:3:1. При этом важно понять, какие классы объединились, и тогда интерпретировать тип взаимодействия.
Необходимо также отметить, что само словосочетание «взаимдействие генов» условно. В действительности взаимодействуют продукты генов, а не сами гены, так что правильнее было бы говорить о взаимодействии фенов, а не о взаимодействии генов. Отсюда понятно, что судить о том, с каким скрещиванием имеет дело экспериментатор: моногибридным, дигибридным или полигибридным - можно только на основании результатов полного гибридологического анализа.
3.3 Полимерия
Наряду с комплементарным и эпистатическим принято также рассматривать взаимодействие генов по типу полимерии. В этом случае разные гены как бы дублируют действие друг друга, и одной доминантной аллели любого из взаимодействующих генов достаточно для проявления изучаемой фенотипической характеристики. Так, при скрещивании растений пастушьей сумки с треугольными плодами (стручками) и с овальными плодами в F1 образуются растения с плодами треугольной формы. При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными стручками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих эднозначно. В этих случаях их обозначают одинаково (А1 и А2). Тогда все генотипы: А1 - А2 -, А1 - а2а2, а2а2А2- будут иметь одинаковую фенотипическую характеристику - треугольные стручки, и только растения а1а1а2а2 будут отличаться - образовывать овальные стручки. Это случай так называемый некумулятивной полимерии.
Однозначные, или полимерные, гены могут действовать и по типу кумулятивной полимерии. Так, шведский генетик Г. Нильсон-Эле в 1908 г. описал серию однозначно действующих генов, которые определяют окраску эндосперма зерен пшеницы. При этом интенсивность окраски зерен оказалась пропорциональной числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3А3, а зерна а1а1а2а2а3а3 не имели пигмента. Между этими крайними типами при расщеплении в F2 наблюдались промежуточные варианты в соотношении 1:6:15:20:15:6:1.