Некоторые последствия использования пестицидов для степных птиц Восточной Европы
Наглядное свидетельство массовой гибели мелких птиц в результате дератизационных работ приводят В.В. Иваницкий и Е.В. Шевченко (1992) на примере монгольского земляного воробья (Pyrgilauda davidiana) в Туве. Там через месяц после обработки чумного очага фосфидом цинка на его территории не удалось обнаружить ни одной птицы, хотя на соседних контрольных участках они оставались вполне обычными, гнездясь с плотностью 5–10 пар/км2.
Здесь, кстати, следует заметить, что поиски погибающих от отравления птиц оказываются, как правило, малоэффективны (Пукинский, 1965; Климов, 1990 и др.), что объясняется, очевидно, быстрой утилизацией трупов различными некрофагами. Последние, как установлено экспериментальными исследованиями в Прикаспийских пустынях, уже в первые сутки собирают более половины (52 %) погибших животных (Семенов, Шейкина, 1946). Поэтому данные о редкой встречаемости павших птиц на обработанных фосфидом цинка территориях (Кондрашкин и др., 1957; Семенов и др., 1957; Абашкин и др., 1971; Климов, 1990 и др.) вряд ли можно считать репрезентативными при оценке воздействия этого пестицида на полезных животных, тем более, что контрольные проверки этих территорий проводились обычно лишь через несколько дней после дератизационных работ (Лисицын и др., 1961; Климов, 1990).
Существенное влияние применения зерновых приманок с фосфидом цинка должно проявляться прежде всего, очевидно, на распространении и численности стадных зерноядных птиц, в частности — журавлей, которые действительно погибают от отравления пестицидами иногда сразу большими стаями (Гусев, 1968; Песков, 1970; Жмуд, 1988 и др.). И, вероятно, депрессия численности журавля-красавки (Anthropoides virgo) в Калмыкии, прослеживаемая по результатам учетов в 1950-е и 1970-е гг. (Банников, 1959; Сурвилло, 1989), была вызвана скорее всего дератизационными работами на этой территории. После приостановки истребительных работ в 1960-е гг., численность журавлей начала здесь, по-видимому, увеличиваться, достигнув к 1970-м гг. 28–39 и даже 70 особей на 100 км автомаршрутов (Голованова, 1982; Сурвилло, 1989), но затем вновь было отмечено ее устойчивое снижение (Сурвилло, 1989а, 1989б). В последующем, после окончательного прекращения массовых противочумных обработок, популяция красавки в Прикаспии быстро восстановилась и, например, в юго-восточных районах Ростовской области, где А.В. Сурвилло отмечал лишь единичных птиц, в 1990 г. гнездилось уже до 5–15 пар/100 км2 (Белик, 1992, 1996б), а в Калмыкии обилие красавки достигло 39 пар/100 км2 (Сотникова, 1991), в два раза превысив уровень общей численности 1972–1975 гг. (Близнюк и др., 1980; Сотникова, 1991).
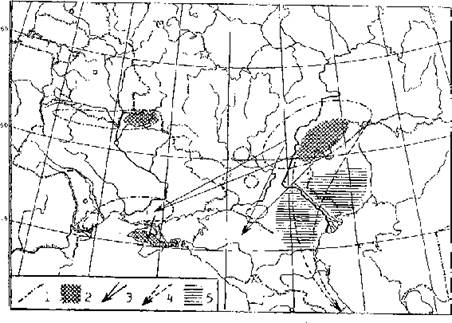
Рис. 6. Структура ареала дрофы в Восточной Европе и основные районы дератизационных работ в Прикаспии: 1 — границы современных гнездовий дрофы, 2 — популяционные ядра важнейших гнездовых группировок, 3 — пути миграций на зимовки, 4 — исчезнувшие миграционные пути, 5 — районы массовой борьбы с грызунами в Прикаспийских очагах чумы.
Высокая смертность от фосфида цинка наблюдается также у дроф (Чуркина, 1964, 1967; Песков, 1970; Федоренко, 1986; Флинт и др., 1992 и др.). И можно предполагать, что в основном именно этот пестицид привел к исчезновению калмыцкой популяции дрофы, до 1960-х гг. являвшейся обычным гнездящимся видом Северо-Западного Прикаспия (Банников, 1959; Близнюк и др., 1980). По всей видимости, с действием этого же фактора связано также и быстрое исчезновение закавказской зимовки дроф, которые до середины 1950-х гг., т. е. до начала применения фосфида цинка, в массе мигрировали через районы интенсивных весенних дератизационных работ (Белоглазов, 1977; Близнюк и др., 1980; Пишванов, 1986; Бутьев и др., 1989). Сохранилась же лишь та их популяция, которая улетала зимовать на запад — в Причерноморье, минуя Прикаспийскую низменность (рис. 6). Однако там зимующие дрофы нередко попадают в “экологическую ловушку”, периодически подвергаясь массовой элиминации во время сильных гололедиц и снегопадов (Бузун, Головач, 1986; Гринченко, 1991 и др.). И поэтому ареал и численность этой популяции дроф, несмотря на все усилия охотоведов, восстанавливается сейчас, в отличие от журавля-красавки, весьма медленно (Флинт и др., 1992).
Кроме зерноядных птиц, от фосфида цинка нередко погибают хищные птицы, подбирающие трупы павших животных (Пукинский, 1965; Климов, 1990), а также насекомоядные птицы, вторичная интоксикация которых связана со сбором жуков и муравьев, объедающих с поверхности зерен растительное масло, используемое в качестве клея для фосфида цинка (Кондрашкин и др., 1957; Климов, 1990). Установлено, например, что муравьи потребляют до 17 % отравленной зерновой приманки (Лисицын и др., 1961), а они сами в пустынно-степных биоценозах являются, в свою очередь, весьма важной составной частью кормового рациона многих видов птиц (Спангенберг, 1946; Волчанецкий и др., 1950; Медведев, Петров, 1959; Бельская, 1965; Рябов, Мосалова, 1966, 1967; Корелов, 1970; Попенко, 1979; Фундукчиев, 1989 и др.). Поэтому не исключено, что дератизационные работы в какой-то степени сказались и на динамике численности некоторых насекомоядных птиц, в частности — степных куликов: кречетки (Chettusia gregaria), большого кроншнепа (Numenius arquata), степной тиркушки (Glareola nordmanni) и др.
Следует отметить, что в силу своей биологической специфики особенно чувствительны к токсинам хищные птицы (Шилова, Переладов, 1974; Ильичев, Галушин, 1978). Очень резкая реакция на фосфид цинка наблюдалась, например, у больших подорликов (Aquila clanga), быстро и почти полностью исчезавших на обработанных этим “родентицидом” территориях в Западной Сибири (табл.; Пукинский, 1965; Данилов, 1976). И сейчас, по-видимому, есть все основания полагать, что происходившая в 1960-е гг. деградация многих популяций большого подорлика (Щербак и др., 1976; Григорьев и др., 1977; Аюпов, 1983; Королькова, 1983; Липсберг, 1983; Лихацкий, 1983 и др.), рекомендованного теперь для включения в Красную книгу России (Мищенко, 1988; Перерва, 1989), была обусловлена в значительной мере воздействием именно фосфида цинка, массово применявшегося с конца 1950-х гг. в туляремийных очагах против водяной полевки (Arvicola terrestris) (Яковлев и др., 1955; Максимов, 1960; Пукинский, 1965; Пукинский, Скалинов, 1967; Данилов, 1976).

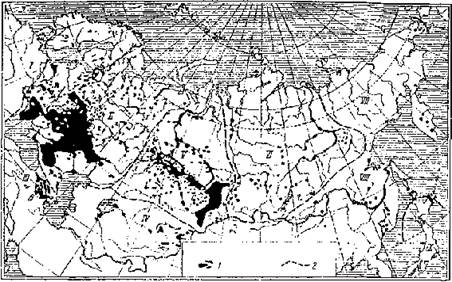
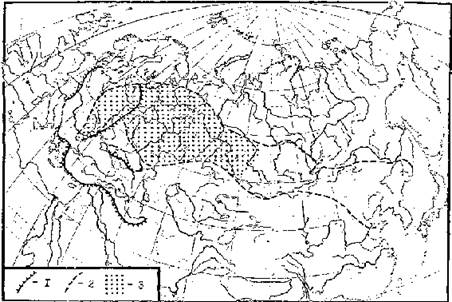
Рис. 7. Гнездовые ареалы подорликов Aquila clanga и A. p. pomarina (внизу), структура ареала водяной полевки (вверху) и распространение природных очагов туляремии в Северной Евразии (посредине) (по: Дементьев, 1951; Неронов, 1965; Олсуфьев, Дунаева, 1970 и др.). Ареалы: 1 — A. p. pomarina, 2 — A. clanga, 3 — исчезающих популяций большого подорлика. Численность водяной полевки: 1 — высокая, 2 — средняя, 3 — низкая; а — стабильная, б — нестабильная. Распространение туляремии: 1 — территории, занятые природными очагами; 2 — границы провинций очаговости.