Страница
4
Гемофилией был поражен царевич Алексей, сын императора России Николая II. Его мать, царица Александра Федоровна, была гетерозиготна по этому аллелю и унаследовала его от своей матери Алисы, которая, в свою очередь, получила его от прабабушки царевича Алексея, английской королевы Виктории:
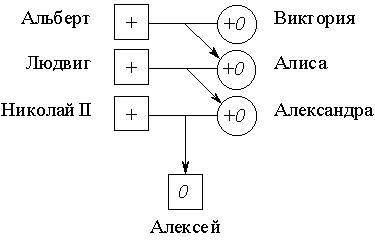
В гетерозиготном состоянии ген гемофилии не проявляется, и поэтому женщины в королевских семьях Европы не страдали гемофилией. Однако многие принцы – потомки королевы Виктории (мутация произошла, по-видимому, именно у нее) получили этот ген и были поражены гемофилией. Вероятность того, что царевич Алексей мог получить дефектный аллель 0 от матери была равна 1/2; с той же вероятностью он мог получить от нее нормальный аллель. Осуществись второе из этих равновероятных событий в образовании гамет, и сценарий судьбы императорской четы выглядел бы иначе.
Рецессивные заболевания, вызываемые генами X-хромосомы, намного реже поражают женщин, чем мужчин, поскольку у них заболевание проявляется только при гомозиготности – наличии рецессивного аллеля в каждой из двух гомологичных X-хромосом; мужчины заболевают во всех случаях, когда их единственная X-хромосома несет дефектный аллель. Количественно это следует из соотношений Харди – Вайнберга Пусть q означает частоту рецессивного аллеля в популяции, т.е. долю X- хромосом, несущих этот аллель. Доля мужчин, имеющих данный аллель и подверженных вызываемой им болезни, равна q. В то же время доля больных женщин равна частоте гомозигот, т.е. q2. Следовательно, число мужчин, больных рецессивным сцепленным с X-хромосомой заболеванием, в 1/q больше, чем число больных женщин. Например, если частота расположенного в X-хромосоме аллеля, вызывающего дальтонизм (неспособность различать цвета), равна 0,05 (т.е. дальтонизм имеется у 5% мужчин), то число дальтоников-мужчин в 20 раз больше, чем дальтоников-женщин.
Пример сцепленного с полом кодоминантного наследования – рыжая окраска домашней кошки, определяемая аллелем у. В гетерозиготном состоянии активными являются оба аллеля (нормальный и у), и поэтому местами шерсть кошки имеет обычную окраску, а местами рыжую. Гомозиготные кошки – целиком рыжие (за исключением возможных белых пятен, вызываемых другим геном, блокирующим синтез пигментов). Самцы же не могут быть частично рыжими; они либо нерыжие, либо целиком рыжие (с возможными белыми пятнами). Исходя из тех же рассуждений, что и абзацем выше, можно было бы заключить, что целиком рыжие коты встречаются гораздо чаще, чем полностью рыжие гомозиготные кошки: частоты их в популяции, соответственно, q и q2, где q – частота «рыжего» аллеля y. Однако в случаях кодоминантного наследования эти рассуждения неприменимы. На самом деле кошки с наличием рыжего цвета (как полностью, так и частично) встречаются гораздо чаще, чем рыжие коты: их частота равна сумме частот гомо- и гетерозигот: q2 + 2q (1– q) = 2q – q2. Например, если частота «рыжего» аллеля равна 0,05, то чисто рыжих кошек должно быть 0,25%, рыжих котов – 5%, а кошек с рыжими пятнами – почти 10%.
Хромосомные перестройки иногда приводят к тому, что фрагмент одной хромосомы «отламывается» и присоединяется к другой хромосоме. Это может произойти и с половыми хромосомами. Так, например, изредка встречаются коты с частично рыжей окраской; это обусловлено тем, что часть X-хромосомы, несущей аллель y, присоединились к Y-хромосоме. В результате по этому локусу наследование осуществляется так же, как и аутосомных генов, т.е. коты с указанным хромосомным нарушением тоже могут быть гетерозиготными, а потому частично рыжими. Однако поломка хромосом приводит к патологиям, в данном случае – глухоте и бесплодию. Это было подмечено давно и выражено во фразе «трехцветные коты глухие». Под третьим цветом здесь понимаются белые пятна. Однако этой патологией страдают и «двуцветные», частично рыжие без белых пятен коты (не путать рыжий цвет с коричневым, который вызывается другим, аутосомным, геном и распространен в ряде пород кошек).
Сцепление с Y-хромосомой. Сведения о генах, находящихся в Y-хромосоме, весьма скудны. Предполагается, что она практически не несет генов, обусловливающих синтез белков, необходимых для функционирования клетки. Но она играет ключевую роль в развитии мужского фенотипа. Отсутствие Y-хромосомы при наличии только одной X-хромосомы приводит к т.н. синдрому Тернера: развитию женского фенотипа с плохо развитыми первичными и вторичными половыми признаками и другими отклонениями от нормы. Встречаются мужчины с добавочной Y-хромосомой (XYY); они высокого роста, агрессивны и нередко аномального поведения. В Y-хромосоме выявлено несколько генов, ответственных за регуляцию синтеза специфических ферментов и гормонов, и нарушения в них приводят к патологиям полового развития. Имеется ряд морфологических признаков, которые, как полагают, определяются генами Y-хромосомы; среди них – развитие волосяного покрова ушей. Подобного рода признаки передаются только по мужской линии: от отца к сыну.
Наследование сложных признаков
Мы рассмотрели правила передачи признака потомству в случае, когда он определяется одним геном. Они справедливы для всех организмов, но тем не менее являются лишь основой для понимания того, как наследуются свойства организма. Дело в том, что многие признаки определяются двумя или большим числом генов. Аллели каждого из этих генов наследуются так, как описано выше. Однако характер наследования признака, который они определяют, зависит от взаимодействия этих аллелей и может быть очень сложным.
Рассмотрим в качестве примера окраску. Окраска шерсти у животных или цветка у растений определяется типом пигмента, его распределением по волосу, перу или лепесткам, пространственным распределением по-разному пигментированных структур и т.д. Все эти частные свойства контролируются разными генами, а в совокупности они все и определяют то, что мы называем окраской.
Например, окраска такого детально изученного экспериментального животного, как мышь, детерминируется по меньшей мере пятью генами. Обычная окраска мыши – серая. Однако сам волос не может быть серым, пигмента такого цвета нет. На самом деле у такой мыши синтезируется и мигрирует в волос черный пигмент, но черная пигментация основания и кончика волоса прерывается желтым кольцом, в котором находится желтый пигмент. Такая окраска называется «агути», именно она и делает мышь «серой». Желтая полоса контролируется геном агути, A, аллель A которого контролирует образование желтой полосы. Рецессивный аллель этого гена, a, блокирует поступление желтого пигмента в волос и обусловливает в гомозиготном состоянии черную окраску мыши. Другой ген, B, контролирует синтез пигментов: доминантный аллель B вызывает образование черного, а рецессивный аллель b коричневого пигмента. В результате, мышь, имеющая оба доминантных аллеля, A и B, – это обычная «серая мышь», а мышь aa и с аллелем B – черная. Однако мышь гомозиготная по второму гену, т.е. bb, и с аллелем A имеет окраску цвета корицы (комбинация коричневого волоса и желтого кольца агути). Мышь же гомозиготная по обоим генам, aabb, полностью коричневая. Имеется ген C, рецессивный аллель которого может прерывать синтез пигментов, и гомозиготная по этому аллелю мышь – белая (альбинос). Ген D контролирует количество пигмента в волосе, так что воспринимаемые глазом различия в интенсивности окраски (например, от светло- до темнокоричневого), определяются различными аллелями этого гена. Ген S детерминирует распределение пигментов по телу и может приводить к пятнистой окраске. Аналогичные гены описаны и у других млекопитающих: домашней кошки, лошади, пушных зверей. Окраска оперения у птиц, надкрылий у жуков, цветка у растений тоже контролируются многими генами; разнообразие сочетаний различных аллелей и определяет то разноцветье, что мы видим в природе.