Страница
2
Латеральное коленчатое тело
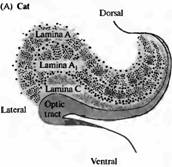
Волокна зрительного нерва начинаются от каждого глаза и заканчиваются на клетках правого и левого латерального коленчатого тела (ЛКТ) (рис. 1), имеющего четко различимую слоистую структуру («коленчатый» — geniculate — означает «изогнутый подобно колену»). В ЛКТ кошки можно увидеть три явных, хорошо различимых слоя клеток (А, А1, С), один из которых (А1) имеет сложное строение и подразделяется далее. У обезьян и других приматов, включая
|
Рис. 1. Латеральное коленчатое тело (ЛКТ). (А) У кошки в ЛКТ имеется три слоя клеток: А, А, и С. (В) ЛКТ обезьяны имеет 6 основных слоев, включающих мелкоклеточные (рагvocellular), или Ρ (3, 4, 5, 6), крупноклеточные (magnocellular), или M (1, 2), разделенные кониоклеточными (koniocellular) слоями (К). У обоих животных каждый слой получает сигналы только от одного глаза и содержит клетки, имеющие специализированные физиологические свойства. |
|
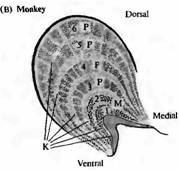
человека, ЛКТ имеет шесть слоев клеток. Клетки в более глубоких слоях 1 и 2 больше по размерам, чем в слоях 3, 4, 5 и 6, из-за чего эти слои и называют соответственно крупноклеточными (M, magnocellular) и мелкоклеточными (Р, parvocellular). Классификация коррелирует также с большими (М) и маленькими (Р) ганглиозными клетками сетчатки, которые посылают свои отростки в ЛКТ. Между каждым M и Р слоями лежит зона очень маленьких клеток: интраламинарный, или кониоклеточный (К, koniocellular) слой. Клетки К слоя отличаются от M и Р клеток по своим функциональным и нейрохимическим свойствам, образуя третий канал информации в зрительную кору.
Как у кошки, так и у обезьяны каждый слой ЛКТ получает сигналы либо от одного, либо от другого глаза. У обезьян слои 6, 4 и 1 получают информацию от контралатерального глаза, а слои 5, 3 и 2 — от ипсилатерального. Разделение хода нервных окончаний от каждого глаза в различные слои было показано при помощи электрофизиологических и целого ряда анатомических методов. Особенно удивительным является тип ветвления отдельного волокна зрительного нерва при инъекции в него фермента пероксидазы хрена (рис. 2).
Образование терминалей ограничено слоями ЛКТ для этого глаза, без выхода за границы этих слоев. Из-за систематического и определенным образом проводимого разделения волокон зрительного нерва в районе хиазмы, все рецептивные поля клеток ЛКТ расположены в зрительном поле противоположной стороны.
|
Рис. 2. Окончания волокон зрительного нерва в ЛКТ кошки. В один из аксонов от зоны с "on" центром контралатерального глаза была введена пероксидаза хрена. Веточки аксона заканчиваются на клетках слоев А и С, но не А1. |
|
|
Рис. 3. Рецептивные поля клеток ШТ. Концентрические рецептивные поля клеток ЛКТ напоминают поля ганглиозных клеток в сетчатке, разделяясь на поля с "on"- и "off''-центром. Показаны ответы клетки с "on"-центром ЛКТ кошки. Полоской над сигналом показана продолжительность освещения. Центральные и периферические зоны нивелируют эффекты друг друга, поэтому диффузное освещение всего рецептивного поля дает только слабые ответы (нижняя запись), еще менее выраженные, чем в ганглиозных клетках сетчатки. |
|


Карты зрительных полей в латеральном коленчатом теле
Важной топографической особенностью является высокая упорядоченность в организации рецептивных полей в пределах каждого слоя ЛКТ. Соседние регионы сетчатки образуют связи с соседними клетками ЛКТ, так что рецептивные поля близрасположенных нейронов ЛКТ перекрываются на большой площади. Клетки центральной зоны сетчатки кошки (регион, где сетчатка кошки имеет маленькие по размеру рецептивные поля с малыми центрами), а также зрительной ямки обезьяны образуют связи с относительно большим количеством клеток в пределах каждого слоя ЛКТ. Подобное же распределение связей было обнаружено и у человека при помощи ЯМР. Число клеток, связанных с периферическими регионами сетчатки, относительно мало. Такая избыточная представленность зрительной ямки отражает высокую плотность фоторецепторов в той зоне, которая необходима для зрения с максимальной остротой. Хотя, наверное, число волокон зрительного нерва и число клеток ЛКТ примерно равны, тем не менее каждый нейрон ЛКТ получает конвергирующие сигналы от нескольких волокон зрительного нерва. Каждое волокно зрительного нерва в свою очередь образует дивергирующие синаптические связи с несколькими нейронами ЛКТ.
Однако каждый слой не только топографически упорядочен, но также и клетки разных слоев находятся в ретинотопическом отношении друг к другу. То есть, если продвигать электрод строго перпендикулярно к поверхности ЛКТ, то сначала будут регистрироваться активность клеток, получающих информацию от соответствующих зон одного, а затем и другого глаза, по мере того, как микроэлектрод пересекает один слой ЛКТ за другим. Расположение рецептивных полей находится в строго соответствующих позициях на обеих сетчатках, т. е. они представляют одну и ту же область зрительного поля. В клетках ЛКТ не происходит значительного смешивания информации от правого и левого глаза и взаимодействия между ними, лишь небольшое количество нейронов (которые имеют рецептивные поля в обоих глазах) возбуждаются исключительно бинокулярно.
Удивительно то, что ответы клеток ЛКТ не имеют разительных отличий от сигналов ганглиозных клеток (рис. 3). Нейроны ЛКТ также имеют концентрически организованные антагонизирующие рецептивные поля, либо с "off"-, либо с "on"-центром, но механизм контраста отрегулирован тоньше, за счет большего соответствия между
|
|
|
|
|
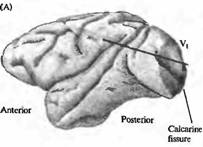
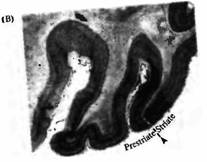
Рис. 4. Связь между первичной зрительной корой (V1) и V2 , V3 , V4 и V5 y обезьян. (А) Плоскость среза проходит через V, и V2 зоны. Четко видна граница между зонами V1 и V2. (В, С) Срез через затылочную кору. На В граница между V1 и V2 показана пунктирной линией в области, где исчезает полосатость. Границы между V2, V3, V4 и V5 зонами выявляются при помощи комбинации физиологических и морфологических исследований. |