Как в клетке возникает разность потенциалов
Оказалось, что при заполнении аксона растворами, содержащими такую же концентрацию К+, как и протоплазма, в аксоне возникал нормальный потенциал покоя порядка 50—60 мВ. При этом выяснилось, что природа отрицательных ионов, которые вводились в волокно вместе с калием, не слишком существенна. Если концентрация калия внутри волокна и в наружной среде была одинаковой, то потенциал покоя в полном соответствии с формулой Нернста не возникал. Если же внутрь волокна накачивали морскую воду, а в наружную среду — раствор с высокой концентрацией калия, то знак потенциала покоя менялся: внутри волокна возникал знак «плюс», а снаружи — «минус»; при этом величина ПП соответствовала формуле Нернста. Таким образом, введение внутрь волокна солевых растворов разного состава подтвердило результаты, полученные ранее при изменении наружной концентрации ионов калия.
Эти опыты показали, во-первых, решающую роль мембраны в возникновении ПП — ведь протоплазма со всеми органеллами и белковыми молекулами попросту отсутствовала, а во-вторых, — решающую роль ионов калия в этом процессе. Мембрана нервного волокна работала так, как это предсказывалось мембранной теорией Бернштейна.
Мембранная теория требует уточнений
Часто, когда теория достигает зрелости, получает признание, живет, начинает расти точность измерений и обнаруживаются отступления от теории, требующие объяснений. Законы Кеплера утверждали, что планеты движутся по эллипсам, но при более точных наблюдениях обнаружилось, что орбиты не вполне эллипсы, что существуют возмущения, которые были объяснены как результат действия на планету силы притяжения других планет. Именно учет возмущений позволил открыть планету Нептун.
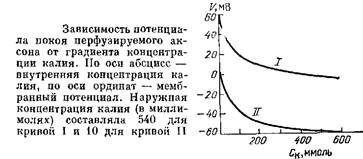
То же самое произошло и с мембранной теорией. Когда методы измерения ПП и ионных концентраций достигли достаточно высокой точности, обнаружилось, что на самом деле измеряемый ПП всегда ниже, чем вычисленный теоретически по формуле Нернста. Ясно, что существовал какой-то «возмущающий» фактор, не учтенный теорией Бернштейна.
Найти причину расхождения эксперимента и теории помог опять-таки кальмар. Работая с гигантским аксоном, А. Ходжкин и Б. Катц установили, что ПП, измеряемый в опыте, оказывается ближе всего к теоретическому в одном особом случае: если из наружной среды убирается натрий.
Но причем здесь натрий? Если, как считал Берн-штейн, мембрана проницаема только для калия, то натрий вообще не должен влиять на ПП! А что если допущение Бернштейна ошибочно? Что будет, если, посягнув на «основу основ» мембранной теории, предположить, что натрий тоже проходит через мембрану? Легко понять, что тогда ионы натрия, которых снаружи клетки гораздо больше, чем внутри, будут идти по градиенту концентрации внутрь, создавая свой нернстовский потенциал, противоположный по знаку тому, который создается калием, и значит, будут снижать ПП. Это равносильно тому, что к мембране подключен не один, а два соединенных навстречу друг другу источника э.д.с. Э.д.с. этих источников определяются концентрационными градиентами калия и натрия. Ходжкин и Катц проверяют свое предположение экспериментально, меняя концентрацию натрия во внешней среде и сравнивая значения ПП с теоретическими. Оказалось, что для совпадения экспериментальных данных с расчетными приходится принять, что сопротивление, через которое идут ионы натрия в 25 раз больше, чем для калия. Это означает, что проницаемость мембраны для ионов натрия, хотя и в 25 раз меньше, чем для калия, но все же не равна нулю.
Вскоре предположение, что ионы натрия могут проходить через мембрану гигантского аксона, было прямо доказано в опытах с радиоактивным изотопом натрия. В ходе этих работ ученые открыли особое вещество, тетродотоксин, блокирующее движение натрия через мембрану. Теперь удалось показать, что если добавить в морскую воду, окружающую гигантский аксон, тетродотоксин и натрий перестает идти через мембрану, то ГШ начинает нарастать и достигает значения, в точности равного тому, которое предсказывается формулой Нернста. Тем самым роль натрия была полностью доказана.
На первый взгляд кажется, что уточнения, внесенные Ходжкином и Катцем в мембранную теорию Бернштейна, несущественны. Всего-то и оказалось, что мембрана пропускает, кроме калия, немного натрия: калий создает потенциал одного знака, натрий — другого, в результате получается ПП чуть ниже, чем только от калия.
Однако на самом деле ситуация меняется в корне.
На мембране устанавливается некоторый промежуточный потенциал, который, естественно, не является равновесным ни для одного из этих ионов. Поэтому и тот и другой ион будут двигаться по градиенту концентрации: ионы натрия — внутрь, а ионы калия — наружу, пока не уравняются концентрации, по крайней мере, одного из них.