Страница
2
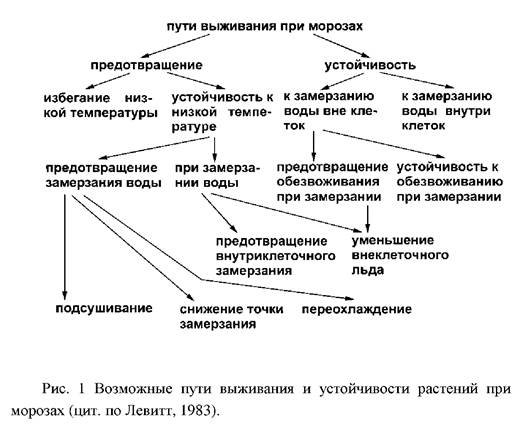
Закаливание озимых растений происходит в три фазы, которые связаны с различными температурными условиями:
– первая фаза закаливания индуцируется снижением температуры до 2 – 5°С, в результате чего исходная морозостойкость повышается на 4–5°С;
– вторая фаза связана с небольшими морозами и в естественных условиях проходит только в том случае, если температура воздуха снижается ниже 0°С;
– третья фаза закаливания может совпадать со второй и зависит от продолжительных морозов, вызывающих обезвоживание клеток.
При изучении процессов закаливания различными исследователями было установлено, что во время первой фазы закаливания в растениях происходят изменения метаболических процессов, таких как гидролиз крахмала и накопление редуцирующих сахаров и водорастворимых белков. Другими метаболическими изменениями, которые тоже являются специфическим ответом растительных тканей на понижение температуры среды, являются превращения липидов и фосфолипидов. Установлено, что низкие положительные температуры повышают ненасыщенность жирных кислот. Было также показано, что содержание фосфолипидов в тканях растений пшеницы возрастало при низкотемпературном закаливании.
Существует ряд доказательств того, что накопление в тканях растений в результате воздействия низких положительных температур некоторых веществ происходит не только вследствие уменьшения их утилизации в ходе ростовых процессов ввиду их приостановки, но и в результате усиления их новообразования. Увеличение ненасыщенности жирных кислот фосфолипидов в условиях пониженных температур, как было показано, тоже в значительно степени связано с их синтезом. Все эти данные свидетельствуют о том, что на первой фазе закаливания у травянистых растений происходит предпочтительны синтез некоторых метаболитов.
Что касается отрицательных температур, которые необходимы для прохождения второй фазы закаливания, то предполагается, что их роль в процессе закаливания не сводится только к физическому действию. По-видимому, она также связана с некоторыми метаболическими процессами, протекающими при этих температурах.
Поскольку прекращение роста растений является необходимым условием прохождения первой фазы закаливания, то метаболические изменения, происходящие в это время, могут быть вызваны изменением гормонального и энергетического баланса.
Принимая во внимание тот факт, что внеклеточное замерзание предотвращает образование внутриклеточного льда, но вызывает при этом обезвоживание макромолекулярных структур клетки, предполагается, что процесс закаливания включает в себя следующие механизмы:
– усиление оттока воды из клетки через мембраны;
– защита клеточных компонентов от действия обезвоживания. Обеспечение оттока воды через мембраны может обеспечиваться путем повышения ненасыщенности липидов. Изменения содержания фосфолипидов также влияют на свойства мембран и повышают их проницаемость для воды, а быстрое снижение содержания фосфолипидов при замораживании вызывает усиленны отток воды в межклетники и защищает клетку от внутриклеточного льдообразования.
При действии мороза повреждения, вызываемые обезвоживанием у закаленных растений, могут быть предотвращены следующими путями:
– структурными и конформационными изменениями компонентов клетки, которые они претерпевают в процессе закаливания;
– защитой компонентов клетки от обезвоживания взаимодействием с низкомолекулярными веществами.
Хотя и установлено, что под действием гипотермии у травянистых растений происходят изменения активности некоторых ферментов, имеется относительно немного экспериментальных доказательств трансформации белковых макромолекул, которые бы вели к повышению их устойчивости к низко температуре. В то же время показано большое значение происходящего во время закаливания растений торможения роста для использования белков, синтезированных в процессе закаливания, на структурную и функциональную реорганизацию клеток.
Исследование процессов холодовой адаптации древесных растений на молекулярном уровне ограничивается наложением на процессы развития холодоустойчивости событий, связанных со входом растения в состояние покоя. R.A. Salzman с соавторами, используя в качестве объекта исследования виноград, создали систему, в которой развитие состояния покоя могло быть индуцировано отдельно от холодовой акклиматизации. С использованием это системы было охарактеризовано дифференциальное накопление ряда белков в почках винограда во время реализации программы нормального входа в состояние покоя совместно с холодовой акклиматизацией и в почках, которые входили только в состояние покоя. Было установлено, что белок с молекулярно массой 47 кДа накапливался в почках винограда во время входа в состояние покоя без холодовой акклиматизации до уровня содержания белка, обнаруженного в находящихся в состоянии покоя и закаленных почках, но не накапливался в закаленных почках, не вошедших в состояние покоя. В то же время 27 кДа LEA-подобный белок накапливался только в закаленных почках. Следовательно, 47 кДа гликопротеин является связанным с состоянием покоя, но не связанным с развитием холодовой акклиматизации, в то время как 27 кДа LEA-подобны белок, по-видимому, более специфичен для холодового закаливания.
Большое значение в регуляции холодо- и морозоустойчивости растений играет абсцизовая кислота. Установлено, что при закаливании растений содержание эндогенно абсцизовой кислоты значительно возрастает, в частности, при закаливании способного к холодовой адаптации вида картофеля Solanum commersonii содержание эндогенной абсцизовой кислоты возрастало в 2,5 раза. У мутанта Arabidopsis thaliana, имеющего низкий уровень содержания эндогенно абсцизовой кислоты, по сравнению с диким типом отсутствовала или была резко снижена способность к холодовой адаптации. В то же время обработка этого мутанта экзогенно абсцизовой кислотой приводила к появлению эффекта адаптации растений к холоду. При этом была показана взаимосвязь между экспрессией регулируемых холодом и регулируемых абсцизовой кислотой генов.
Аналогичные результаты были получены и при изучении других видов растений. Было проведено определение морозоустойчивости побегов, корней и тканей эпикотиля гороха сорта Alaska двух генотипов: дефицитного по содержанию абсцизовой кислоты мутанта «wil» и его дикого типа при различных типах стресса. В ходе исследований спектры белков изучались при помощи двумерного SDS-PAGE электрофореза. При этом было установлено, что холодовая обработка индуцировала образование семи белков в побегах, трех – в эпикотиле и двух – в корнях гороха. В тканях побегов пять из семи новых белков накапливались также в ответ на обработку абсцизовой кислотой. Полипептид с молекулярной массой 24 кДа продуцировался и в мутантных, и в «диких» проростках и тканях эпикотиля только после холодовой обработки.
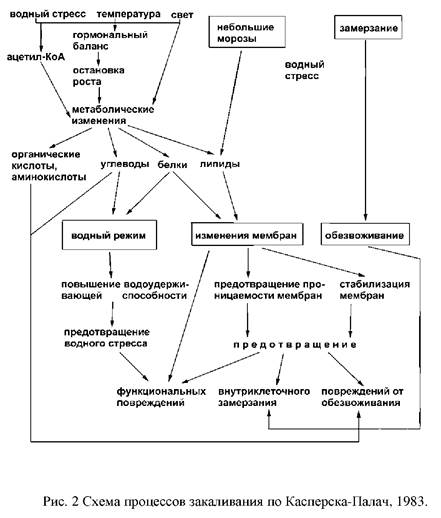
Таким образом, существенным этапом перехода от стрессовых к адаптационным реакциям является изменение экспрессии генов, выражающееся в ингибировании активных генов, в норме контролирующих рост, развитие и фотосинтез. При этом активируется система генов контроля за устойчивостью: происходит синтез новых белков, специфических адаптогенов и стресс-протекторов. Завершается эта перестройка структурными изменениями в организме растения.