Страница
5
2.4 Молекулярная характеристика гена FIM (львиный зев)
Ген львиного зева FIM был клонирован с использованием метода транспозоновой мишени. Мутанты fun 619 и fim 620 с инсерцией мобильных элементов были получены в описанном выше эксперименте по мутагенезу. Ген арабидопсиса UFO был клонирован по гомологии с геном FIM: фрагмент гена FIM почти полной длины использовали как пробу для гибридизациии с геномной библиотекой арабидопсиса. Нуклеотидные последовательности генов FIM и UFO обладают значительной степенью гомологии. В то же время последовательности этих генов не обнаруживают значительного сходства с какими-либо другими нуклеотидными последовательностями, имеющимися в базах данных. Оба гена, FIM и UFO, активируются в молодых цветковых меристемах раньше генов типа органов цветка. Инициация их экспрессии наблюдается в центре цветковой меристемы, и затем эта область «расходится» в стороны и превращается в кольцо, в центре которого РНК FIM или UFO уже не обнаруживаются. Еще позднее экспрессия этих генов ограничивается небольшой областью между мутовками лепестков и других органов. UFO работает только в области между чашелистиками и лепестками, в то время как ген FIM экспресси-руется как между мутовками чашелистиков и лепестков, так и между мутовками лепестков и тычинок.
Между гомологами FIM/UFO, как и в случае другой пары FLO/LFY, обнаруживаются и другие различия. У цветков мутантов ufo, как и у мутантов Ify, образуются прицветники. Таким образом, одной из функций генов LFY и UFO является подавление образования прицветников. Подобной функцией не обладают их гомологи FLO и FIM. Ген UFO начинает действовать чуть раньше, чем ген FIM. Транскрипты UFO обнаруживаются на более ранней стадии, чем транскрипты FIM.
Мутации ufo выражаются в изменении размеров, количества и закладки органов цветка, а также в их слиянии. Это говорит о том, что ген UFO участвует в регуляции клеточных делений и пролиферации органов. Подтверждается это и фенотипом трансгенных растений арабидопсиса с геном UFO, слитым с конститутивным промотором p35S ВМЦК: в цветках таких растений развивается больше лепестков и тычинок, при этом общее количество лепестков и тычинок может быть как больше, так и меньше восьми, а количество репродуктивных органов может достигать шестнадцати (против шести в цветках дикого типа). Гены UFO/FIM характеризуются наличием F-бокса, который имеется также в генах, контролирующих синтез таких белков, как циклин А, циклин F и Cdc4, вовлеченных в контроль клеточного цикла у дрожжей. Некоторые белки с F-боксом, например MetSop и GRR1 дрожжей, функционируют как ингибиторы транскрипции. Белок Fim связывается с белками семейства Fap (от EIM-associated £roteins), которые очень напоминают белки Skpl дрожжей и человека. Последние образуют комплексы с белками, содержащими F-бокс, вызывая деградацию белков и развитие клеточного цикла. Ген FIM играет также ключевую роль в активации генов идентичности цветковой меристемы.
2.5 Молекулярная характеристика генов CEN (львиный зев) и TFL1 (арабидопсис)
Гены CEN и TFL1 активируются в зрелых соцветиях дикого типа чуть ниже верхней части апикальной меристемы соцветия (рис. 3). РНК TFL1 также обнаруживается в латеральных генеративных меристемах и по всей длине генеративного стебля. Таким образом, вероятно, работа гена TFL1 в генеративном стебле приводит к задержке перехода к цветению. Ген CEN экспрессируется в генеративном стебле очень слабо. Вероятно, поэтому мутации сеп, в отличие от мутаций tfl, не влияют на время перехода к цветению. Экспрессия гена TFL1 в апикальной меристеме ингиби-рует работу в ней генов LFY и API, а гена CEN ингибирует экспрессию гена FLO и таким образом предотвращает развитие апекса побега как цветочной меристемы (рис. 11.34). Неожиданным результатом анализа взаимодействия CEN с другими генами оказалось то, что ген FLO в свою очередь необходим для экспрессии CEN в апикальной меристеме. В мутантных по FLO растениях, у которых формируется нефункциональная РНК FLO, РНК CEN не обнаруживается.
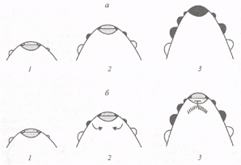
Рис. 3. Схематическое изображение начальных этапов развития генеративной меристемы мутанта сеп (а) и растений львиного зева дикого типа (б).
1 - момент закладки цветковых меристем; 2 и 3 - апекс генеративной меристемы через 2 и 6 дней с момента закладки цветковых меристем. Цветковые меристемы показаны черным цветом, вегетативные - белым, апикальные меристемы - точками. Прицветники не показаны. В мутанте cen ген FLO после некоторой задержки активируется в апикальной меристеме. В растениях дикого типа экспрессия FLO индуцирует экспрессию CEN. В субапикальной зоне (сигнал от FLO показан стрелками, зона экспрессии GEN заштрихована). Активация CEN ингибирует экспрессию FLO в апикальной меристеме.
Вывод
Гены, о которых говорилось выше, необходимы для успешного перехода к цветению. Можно выделить две группы таких генов: гены, влияющие на время индукции цветения, и гены идентичности цветковых меристем.