Страница
2
2. Вероятность образования новых столонов в колонии (Р) вычисляли делением числа колоний, в которых при заданных условиях кормления образовался хотя бы один столон, на общее число колоний, содержащихся при таких же условиях. Вероятность выражали в процентах.
3. Разветвленность гидроризы (B—2/L) показывает, сколько боковых столонов приходится на единицу длины гидроризы колонии. Для вычисления этого параметра число верхушек столонов без двух делили на суммарную длину столонов колонии. В неразветвленной колонии, которая состоит из одного столона с двумя верхушками (В =2), этот показатель принимает нулевое значение.

Рис. 3 . . Рис. 4
Рис. 3. Изменение скорости роста столона (/) и коэффициента вариации (С. V.} (2) с течением времени (сут). Каждая точка соответствует среднему значению для выборки из 10 колоний
Рис. 4. Зависимость средней скорости роста верхушки столона {l/B) от интенсивности питания колонии (F/Lt). 1—значения для неветвящихся колоний; 2—значения накануне ветвления колонии. Прямая определяется уравнением l/B = 0,Ol5 F/lt —0,095, коэффициенты которого вычислены по методу наименьших квадратов для интервала 0 < F/Lt £ O,150. Распределение экспериментальных точек на указанном интервале достоверно соответствует уравнению прямой (коэффициент корреляции С. V. = 0,0664).
4. Удельное число гидрантов, на рост которых расходуется питательный материал (Hg2/L=Hi2—Hd /L.). В литературе имеются указания на то, что клеточный материал, освобождающийся при рассасывании гидрантов, может переноситься гидроплазмой в другие части колонии и использоваться там для их роста (Nathanson, 1955; Hale, 1964; Braverman, 1974). Поэтому мы предположили, что количество питательного материала, поступающего в колонию в результате дедифференцировки одного старого гидранта, примерно эквивалентно тому количеству материала, которое расходуется на рост одного нового.
Для вычисления введенного показателя разность между числом гидрантов, находящихся в процессе формирования (такими мы считали растущие гидранты, у которых было менее шести щупалец), и числом дегенерирующих делили на суммарную длину гидроризы колонии.
5. Объем ткани единицы длины ценосарка (v/L) вычисляли, измеряя диаметр ценосарка и гастроваскулярной полости, и затем по соответствующей формуле—площадь сечения стенки ценосарка в данном месте столона. Сделав большое количество замеров в колонии, вычисляли среднюю площадь, сечения ценосарка в ней. Объем ткани единицы длины ценосарка имеет то же численное значение и ту же размерность (мм), что и площадь сечения.
Измерение скоростей роста столонов в 10 колониях, получивших примерно одинаковое небольшое количество пищи,—0,126 (С. V.=0,191) науплиусов/(мм×сут), показало, что в первые 6 сут после выделения из материнской колонии рост столона еще не стабилизирован (рис. 3). В этот период изменение скорости роста носит нерегулярный характер и вариабельность средних ее значений в разных колониях непостоянна в течение времени, но, начиная с 7-х сут, стабилизируется. По этой причине мы рассматривали лишь те морфологические показатели, которые относились к периоду стабилизированного роста колоний — с 7-х по 14-е сут.
Когда интенсивность питания не превышает величины 0,200 науплиусов/(мм×сут), средняя скорость роста столона тем выше, чем больше среднее количество пищи, полученное колонией (рис. 4). Распределение экспериментальных точек достоверно соответствует уравнению линейной регрессии l/B = 0,015F/t - 0,095 (г-0,664; n - 36), вычисленному по методу наименьших квадратов. При более интенсивном питании ускорения роста не происходит. В этих условиях уже в большинстве колоний (более 55%) образуются новые столоны, за счет которых об щий прирост гидроризы продолжает увеличиваться (рис. 5), несмотря на то, что скорость роста каждого отдельного столона больше не возрастает.
На характер ветвления столонов влияют количество пищи и динамика поступления ее в колонию. Кормление проводили двумя разными способами. В одной группе (33 колонии) удельное среднесуточное количество пищи было в каждой колонии довольно постоянно. Средний коэффициент вариации показателя интенсивности питания (F/Lt) для 23 колоний (n =23) составил 0,37. В другой группе (33 колонии) каждая колония сначала получала ежедневно одно и то же небольшое количество пищи (F/Lt=0,137; C.V.=0,55) и новых столонов в них не образовывалось. На 7—14-е сут роста каждая колония получала одноразовую избыточную дозу корма (F/L = 0,434; С. V. = 0,32), так что среднее значение показателя F/Lt изменялось в среднем на 0,038 науплиусов/(мм×сут) (C.V.=0,57). Такое “импульсное” кормление приводило к образованию боковых столонов. Условия ветвления столонов в первой и второй группах колоний несколько отличались.
В первой группе вероятность образования почки столона (Р) возрастает с увеличением интенсивности питания (рис. 6, а). При интенсивности питания менее 0,100 науплиусов/(мм×сут) новых столонов не образуется. Далее при увеличении количества пищи от 0,100 до 0,250 науплиусов/(мм×сут) вероятность образования столонов увеличивается и при интенсивности питания 0,250 и выше составляет 100%. Среднее число образующихся почек столонов почти не зависит от интенсивности питания и составляет 3,5—4 почки на колонию (рис. 6, б). Однако показатель разветвленности гидроризы B—2/L при этом все же несколько увеличивается (рис. 6, в). Это связано с тем, что при обильном кормлении ветвление в колонии начинается раньше, когда длина гидроризы еще невелика (рис.6,г). При более умеренном питании ветвление начинается на 2-3 дня позже, когда размер колонии несколько больше.
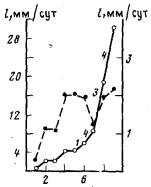
Рис. 5. Изменение суточного прироста гидроризы (l) (сплошная линия) и средней скорости роста верхушек столонов (1/В) (пунктирная линия) с течением времени. Цифры рядом с отрезками сплошной линии означают число почек столонов, образовавшихся за сутки
Импульсное кормление вызывает более или менее значительное увеличение средней интенсивности питания, в среднем на 0,038 науплиусов/(мм×сут) (С.V.=0,56; n=30). Оказалось, что ни вероятность ветвления гидроризы, ни число образующихся столонов практически не зависят от величины интенсивности питания, возросшей в результате импульсного кормления (рис. 6, д, е). Вероятность ветвления возрастает от 70 до 80% при увеличении показателя F/Lt от 0,100 до 0,250 науплиусов/(мм×сут). Среднее число образующихся столонов при этом возрастает от 3,15 до 3,30 экз/колонию. Такое изменение не является достоверным (t=0,200; п=28). Почки столонов появляются через 26±2 ч после импульсного кормления.
В тех колониях, которые получали мало пищи (менее 0,100 науплиусов/(мм×сут), формирующихся гидрантов было меньше, чем рассасывающихся. В этих случаях показатель Hg/L принимает, отрицательные значения (рис. 7). При увеличении интенсивности питания величина этого показателя возрастает, причем какого-либо предельного его значения в наших экспериментах обнаружено не было. Среднее значение параметра Hg/L (0,123) достоверно выше (t=2,227; n=24) в тех колониях, где не образовалось ни одного бокового столона, и ниже (Hg/L=0,056) в тех, которые ветвятся.