Биосинтез антител. Структура и специфичность антигенов
Антигенная специфичность гаптенов зависит не только от их химической структуры, но и от способа пришивки к белку-носителю, в частности от того, какая функциональная группа гаптена была использована для конъюгирования. Часто, при получении конъюгатов для иммунизации гаптены пришивают не непосредственно к молекуле белка-носителя, а через пространственную «ножку», содержащую обычно 4—6 углеродных атомов. В этом случае сама «ножка» в комплексе с гаптеном выступает в качестве составной антигенной детерминанты и образующиеся антитела могут обладать меньшей эффективностью связывания с нативным гаптеном, чем с таким связанным через «ножку» гаптеном.
В некоторых видах иммуноферментного анализа в качестве одного из реагентов используют гаптен, меченный ферментом. Если связывание в таком конъюгате аналогично способу пришивки в конъюгате гаптена с белком-иосителем для получения антител, то говорят, что в анализе используют гомологичные антитела. Если же структура «ножки» и способ пришивки гаптена в обоих случаях различны, то говорят о гетерологичных антителах. Применение того или иного вида антител или конъюгатов в иммуноферментном анализе может весьма сильно сказываться на его чувствительности и некоторых других характеристиках.
Весьма существенным фактором для специфичности является химическая структура «нодаскы», в частности ее длина и ближайшее окружение гаптена. Все эти моменты крайне важно учитывать при разработке методов иммунохимического анализа гаптенов.
Сильное влияние различных факторов на антигенную структуру и специфичность гаптенов, по-видимому, объясняется их ограниченными размерами и особенностями структуры активных центров антител.
Знание антигенной структуры и специфичности гаптенов имеет важное значение для создания методов иммунохимического определения различных физиологически активных соединений, так как многие из них претерпевают различные биохимические превращения, в результате чего образуется группа близкородственных метаболитов.
3. Общая структурная характеристика молекул иммуноглобулинов
Антитела в организме выполняют две основные функции. Первая — это распознавание и специфическое связывание соответствующих антигенов, вторая — эффекторная, заключающаяся в индукции важнейших физиологических процессов, направленных на уничтожение антигена: лизис чужеродных клеток через активацию системы комплемента, стимуляция специализированных иммунокомпетентных клеток, выделение фармакологически активных веществ и т.д.
Развитие иммунохимии в течение последних 25 лет позволило установить строение антител, выявить стереохимические основы их функционирования. Особое внимание было уделено изучению структуры активных центров антител, что привело к созданию полицентровой модели связывания антигена. Исследования динамических структурных свойств иммуноглобулинов способствовало установлению характера связи между антигенсвязывающими и эффекторными функциями.
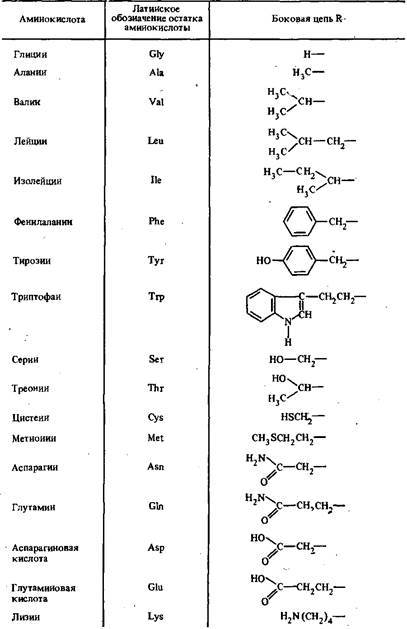
Иммуноглобулины по своей химической структуре относятся к большому классу природных соединений — гликопротеидам, высокомолекулярным соединениям, состоящим из последовательности L-аминокислот, соединенных между собой пептидными связями.
Отдельные аминокислоты отличаются между собой боковыми заместителями R. В состав белков, в том числе иммуноглобулинов, входят двадцать аминокислотных остатков.
Полипептидная цепочка за счет образования водородных связей между карбонильным атомом кислорода и атомом водорода аминогрупп отдельных аминокислотных остатков способна определенным образом укладываться в пространстве, образуя так называемые а-спиральные участки и структуру. Такая локальная упорядоченная конформация отдельных участков полипептидной цепи получила название вторичной структуры.
В целом вся полипептидная цепь образует компактную трехмерную структуру – третичную структуру: В одном растворе молекула белка сворачивается так, чтобы неполярные, или гидрофобные, боковые цепи аминокислотных остатков находились во внутренней, малодоступной для молекул воды области, а полярные, или ионизированные, группы образовывали внешний контактирующий с водой слой. Такое расположение аминокислотных остатков полипептидной цепи является термодинамическим наиболее выгодным состоянием, причем следует отметить, что это сворачивание пептидной цепи является высокоспецифичным и обусловлено первичной структурой молекулы.
Помимо рассмотренных причин сворачивания белков, обусловленных так называемыми гидрофобными взаимодействиями, определенный вклад в' стабилизацию трехмерной структуры вносят дисперсионные силы Лондона, возникающие в результате комплементарного распределения электронных облаков отдельных, рядом расположенных атомов. Энергия этого типа взаимодействий сильно зависит от расстояния между атомами и максимальна при так называемом ван-дер-ваальсовом расстоянии контакта, равном сумме ван-дер-ваальсовых радиусов взаимодействующих атомов.
Дополнительный вклад в поддержание трехмерной структуры молекул белков дают водородные и электростатические связи между боковыми группами аминокислот, а также ковалентные связи между отдельными частями полипептидной цепочки, например дисульфидные, или S—S-связи, возникающие между двумя остатками цистеина.
Некоторые молекулы белков состоят из несвязанных между собой ковалентно отдельных субъединиц. Такая пространственная организация получила название четвертичной структуры белков.
Несмотря на огромное разнообразие антител и их гетерогенность, все они обладают некоторыми общими структурными элементами, обеспечивающими выполнение их основных функций.
По своим антигенным, эффекторным свойствам и структурным особенностям иммуноглобулины подразделяются на пять основных классов: IgM, IgG, IgA, IgD и IgE.
Таблица 1 - Основные аминокислоты, входящие в состав белков
Общей структурной единицей всех иммуноглобулинов является комплекс из четырех полипептидных цепей — двух идентичных между собой легких цепей с молекулярной массой 23000 каждая и тяжелый с молекулярной массой по 53000.
Каждая из лёгких цепей прочно соединена с концевыми участками тяжелых цепей благодаря наличию межцепочечных ди-сульфидных связей и множеству слабых гидрофобных, электростатических и других межатомных взаимодействий. Аналогичные связи существуют и между свободными участками тяжелых цепей. В целом структура такого комплекса напоминает латинскую букву «V» или «Т» и характерна для иммуноглобулинов классов IgG, IgD и IgE.
При действии протеолитического фермента папаина молекула IgG распадается на три фрагмента, два из которых идентичны и сохраняют способность связывать антигены и третий, способный к кристаллизации. Именно Fc-фрагмент ответствен за эффекторную функцию антител — связывание белка комплемента Clq, транспорт через мембраны, взаимодействие с мембранными рецепторами и т.д.
Другой протеолитический фермент пепсин "разрывает пептидную связь, расположенную ближе к СООН-концу цепи, от S—S-связи между Н-цепями в Fc-фрагменте. В результате образуются так называемый pFc-фрагмент, представляющий собой остатки тяжелых цепей, и соединенные дисульфидными связями два Fа-фрагмента, обозначаемые как ¥2-фрагмент. Последний также сохраняет способность к связыванию антигенов.