Аэробное и анаэробное дыхание растений
2) цикл трикарбоновых кислот (цикл Кребса);
3) заключительная стадия окисления – электронтранспортная цепь.
На первой стадии пировиноградная кислота взаимодействует с веществом, которое называют коферментом А, в результате чего образуется ацетилкофермент а с высокоэнергетической связью. При этом от молекулы пировиноградной кислоты отщепляется молекула CO![]() (первая) и атомы водорода, которые запасаются в форме НАД ∙ Н + Н
(первая) и атомы водорода, которые запасаются в форме НАД ∙ Н + Н![]() .
.
Вторая стадия – цикл Кребса (рис. 1)

В цикл Кребса вступает ацетил–КоА, образованный на предыдущей стадии. Ацетил–КоА взаимодействует со щавелево-уксусной кислотой, в результате образуется шестиуглеродная лимонная кислота. Для этой реакции требуется энергия; ее поставляет высокоэнергетическая связь ацетил–КоА. В конце цикла щавелево-лимонная кислота регенерируется в прежнем виде. Теперь она способна вступить в реакцию с новой молекулой ацетил–КоА, и цикл повторяется. Суммарно реакция цикла может быть выражена следующим уравнением:
ацетил-КоА + 3H![]() O + 3НАД
O + 3НАД![]() + ФАД + АДФ + Н
+ ФАД + АДФ + Н![]() РО
РО![]() →
→
КоА + 2CO![]() + 3НАД ∙ Н + Н
+ 3НАД ∙ Н + Н![]() +ФАД ∙ H
+ФАД ∙ H![]() + АТФ.
+ АТФ.
Таким образом, в результате распада одной молекулы пировиноградной кислоты в аэробной фазе (декарбоксилирование ПВК и цикла Кребса) выделяется 3CO![]() , 4 НАД ∙ Н + Н
, 4 НАД ∙ Н + Н![]() , ФАД ∙ H
, ФАД ∙ H![]() . Суммарно реакцию гликолиза, окислительного декарбоксилирования и цикла Кребса можно записать в следующем виде:
. Суммарно реакцию гликолиза, окислительного декарбоксилирования и цикла Кребса можно записать в следующем виде:
C![]() H
H![]() O
O![]() + 6 H
+ 6 H![]() O + 10 НАД + 2ФАД →
O + 10 НАД + 2ФАД →
6CO![]() + 4АТФ + 10 НАД ∙ Н + Н
+ 4АТФ + 10 НАД ∙ Н + Н![]() + 2ФАД ∙ H
+ 2ФАД ∙ H![]() .
.
Третья стадия – электротранспортная цепь.
Пары водородных атомов, отщепляемые от промежуточных продуктов в реакциях дегидрирования при гликолизе и в цикле Кребса, в конце концов, окисляются молекулярным кислородом до H![]() O с одновременным фосфолированием АДФ в АТФ. Происходит это тогда, когда водород, отделившийся от НАД ∙ H
O с одновременным фосфолированием АДФ в АТФ. Происходит это тогда, когда водород, отделившийся от НАД ∙ H![]() и ФАД ∙ H
и ФАД ∙ H![]() , передается по цепи переносчиков, встроенных во внутреннюю мембрану митохондрий. Пары атомов водорода 2Н можно рассматривать как 2 Н
, передается по цепи переносчиков, встроенных во внутреннюю мембрану митохондрий. Пары атомов водорода 2Н можно рассматривать как 2 Н![]() + 2е
+ 2е![]() . Движущей силой транспорта атомов водорода в дыхательной цепи является разность потенциалов.
. Движущей силой транспорта атомов водорода в дыхательной цепи является разность потенциалов.
С помощью переносчиков ионы водорода Н![]() переносятся с внутренней стороны мембраны на ее внешнюю сторону, иначе говоря, из матрикса митохондрии в межмембранное пространство (рис. 2).
переносятся с внутренней стороны мембраны на ее внешнюю сторону, иначе говоря, из матрикса митохондрии в межмембранное пространство (рис. 2).
|
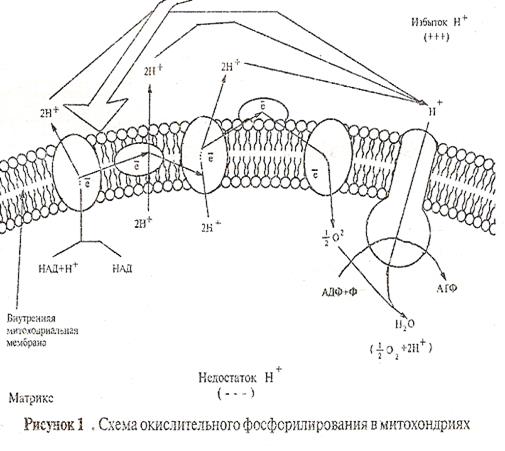
При переносе пары электронов от над на кислород они пересекают мембрану три раза, и этот процесс сопровождается выделением на внешнюю сторону мембраны шести протонов. На заключительном этапе протоны переносятся на внутреннюю сторону мембраны и акцептируются кислородом:
½ O![]() + 2е
+ 2е![]() → O
→ O![]() .
.
В результате такого переноса ионов Н![]() на внешнюю сторону мембраны митохондрий в перимитохондриальном пространстве создается концентрация их, т.е. возникает электрохимический градиент протонов .
на внешнюю сторону мембраны митохондрий в перимитохондриальном пространстве создается концентрация их, т.е. возникает электрохимический градиент протонов .
Когда протонный градиент достигает определенной величины, ионы водорода из Н![]() -резервуара движутся по специальным каналам в мембране, и их запас энергии используется для синтеза АТФ. В матриксе они соединяются с заряженными частичками О
-резервуара движутся по специальным каналам в мембране, и их запас энергии используется для синтеза АТФ. В матриксе они соединяются с заряженными частичками О![]() , и образуется вода: 2Н
, и образуется вода: 2Н![]() + О²ˉ → H
+ О²ˉ → H![]() O.
O.
1.1 Окислительное фосфолирование
Процесс образования АТФ в результате переноса ионов Н![]() через мембрану митохондрии получил название окислительного фосфолирования. Он осуществляется при участии фермента АТФ-синтетазы. Молекулы АТФ-синтетазы располагаются в виде сферических гранул на внутренней стороне внутренней мембраны митохондрий.
через мембрану митохондрии получил название окислительного фосфолирования. Он осуществляется при участии фермента АТФ-синтетазы. Молекулы АТФ-синтетазы располагаются в виде сферических гранул на внутренней стороне внутренней мембраны митохондрий.
В результате расщепления двух молекул пировиноградной кислоты и переноса ионов водорода через мембрану по специальным каналам синтезируется в целом 36 молекул АТФ (2 молекулы в цикле Кребса и 34 молекулы в результате переноса ионов Н![]() через мембрану).
через мембрану).
Суммарное уравнение аэробного дыхания можно выразить следующим образом:
C![]() H
H![]() O
O![]() + O
+ O![]() + 6H
+ 6H![]() O + 38АДФ + 38Н
O + 38АДФ + 38Н![]() РО
РО![]() →
→
6CO![]() + 12H
+ 12H![]() O + 38АТФ
O + 38АТФ