Нейроподобный элемент (нейрон)
2.1.2. Сети с симметричными связями.
Ансамблевые нейронные сети. Минский и Пейперт отмечали, что недостатки простых персептронов можно преодолеть как с помощью многослойных сетей (см. выше), так и введением в сеть обратных связей, допускающих циркуляцию сигналов по замкнутым контурам. Использовать свойства такого рода сетей для моделирования функций мозга еще в 1949 г. предложил Хебб.
Согласно взглядам Хебба нервные клетки мозга соединены друг с другом большим количеством прямых и обратных возбуждающих связей и образуют нейронную сеть. Каждый нейрон осуществляет пространственно-временную суммацию приходящих к нему сигналов от возбуждающих, определяя потенциал на своей мембране. Когда потенциал на мембране превышает пороговое значение, нейрон возбуждается. Нейрон обладает рефрактерностью и усталостью. Эффективность связей может изменяться в процессе функционирования сети, повышаясь между одновременно возбужденными нейронами. Это приводит к объединению нейронов в клеточные ансамбли - группы клеток, которые чаще всего возбуждались вместе, и к обособлению ансамблей друг от друга. При возбуждении достаточной части ансамбля он возбуждается целиком. Различные ансамбли могут пересекаться: один и тот же нейрон может входить в разные ансамбли. Электрическая активность мозга обусловлена последовательным возбуждением отдельных ансамблей.
Идеи Хебба оказали большое воздействие на представления о работе мозга и послужили основой для создания нейронных моделей долговременной памяти. Действительно, ансамблевую нейронную сеть можно рассматривать как структуру, реализующую функции распределенной ассоциативной памяти. Формирование ансамблей в такой сети соответствует запоминанию образов (признаков, объектов, событий, понятий), закодированных паттерном активности нейронов, а сформированные ансамбли являются их внутренним представлением. Процесс возбуждения всего ансамбля при активации части его нейронов можно интерпретировать как извлечение запомненной информации по ее части - ключу памяти.
Модель памяти на основе ансамблевой нейронной сети обладает некоторыми свойствами, присущими биологической памяти, таким, как ассоциативность, распределенность, параллельность, устойчивость к шуму и сбоям, надежность. Проводятся также структурные аналоги между ансамблевыми моделями нейронных сетей и строением коры головного мозга. Имеются экспериментальные данные о синаптической пластичности, постулированной Хеббом.
Модель ансамблиевой сети состоит из большого количества нейронов, каждый из которых обычно соединен со всеми другими элементами сети. Входной образ подается на сеть путем активации нужных нейроподобных элементов. Обучение ансамблиевой сети описано в параграфе .
Сеть Хопфилда. Хотя многочисленные результаты моделирования демонстрировали стабильность ансамблевых сетей с обратными связями и хеббовским правилом обучения (эволюцию сети к устойчивому состоянию), отсутствие математического обоснавания такого поведения препятствовало их популярности.
В 1982 г. американский биофизик Джон Хопвилд опубликовал статью, где поведение модели полносвязной сети бинарных нейроподобных элементов с симметричными связями (wij = wji). элементы функционировали в асинхронном режиме, т.е. каждый нейрон в случайные моменты времени с некоторой средней частотой определял свое состояние с правилом (1.3). Это позволило описать поведение сети ка релаксационный процесс, при котором минимизируется функция (гамильтониан) модели.
Машина Больцмана представляет собой стохастический вариант сети Хопфилда. Бинарные нейроподобные элементы (блоки) трактуются здесь представители элементарных гипотез, а веса - как слабые парные взаимоограничения между ними. Положительный вес связи указывает, что две гипотезы стремятся поддерживать друг друга, а отрицательный - на их несовместимость. Симметрия связей позволяет проанализировать поведение сети с использованием энергетической функции (гамильтониана). Энергию определенного паттерна активности можно интерпретировать как степень нарушения ограничений, присутствующих в проблемной области, со стороны конкретной комбинации гипотез или как стоимостную функцию, которая должна быть минимизирована для решения оптимизационной задачи. Существует возможность, однако, попадания сети в локальный мимнимум, что крайне не желательно для оптимизационных задач. Чтобы сеть могла выбраться из локального энергетического минимума, в машине Больцмана применяется вероятностное правило срабатывания блоков:
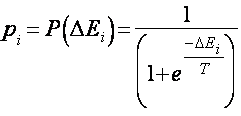 , (2.1)
, (2.1)
где pi - вероятность нахождения i-го блока в единичном состоянии;
P (x) - сигмоидная функция (рис. 1.2. б);
T - параметр, аналогичный температуре. При T®0 это правило переходит в правило срабатывания детерминированных элементов (1.3), а при повышении температуры увеличивается вероятность перехода системы в состояние с большей энергией.
Обучение машины Больцмана описано в параграфе 2.2.
2.1.3. Сети с латеральным торможением.
Карты признаков Кохонена. Обычно в качестве входных образов в моделях ассоциативной памяти используются некоторые внутренние представления сенсорной информации, прошедшей, как считается, необходимую предобработку. Один из нейросетевых вариантов такой переработки предложен Кохоненом. Его алгоритм формирует одно- или двумерную карту «карту» признаков путем нелинейного «сплющивания» многомерного сигнального пространства. При этом предполагается, что такое отображение должно сохранять топологические отношения, существующие между входными сигналами.
Структура нейронной сети, в которой реализуется формирование карт признаков, приведена на рис. 2.1. Нейроны, имеющие сигмоидную характеристику, расположены в виде одно- и двумерного слоя слоя по аналогии со слоистым строением коры. На каждый нейрон поступают два вида связей: mij, которые интерпретируются как связи от сенсорных входов или из других областей, и wjk - латеральные связи от нейронов одного слоя, характер которых зависит от расстояния между нейронами. Функция взаимодействия нейронов одного слоя имеет вид «мексиканской шляпы» (рис. 2.2.), что соответствует некоторым нейробиологическим данным. Близко расположенные нейроны возбуждают друг друга, с увеличением расстояния возбуждение сменяется торможением, а затем опять появляются слабые возбуждающие связи, которые по-видимому, выполняют ассоциативные функции и в данной модели не используются.

Эффект наличия латеральных связей с радиусом действия порядка размеров сети проявляется в следующем. Если на каждый нейрон подать (например, через связи от сенсорных входов mij) имеющий небольшой максимум случайный сигнал Si, то в процессе релаксации сети осуществляется повышение его контрасности. В результате вокруг первоначального максимума образуется «пузырек» выходной активности нейронов (рис. 2.3.).